У всіх живих клітинах білки синтезуються рибосомами . Рибосома є великою макромолекулою зі складною асиметричною четвертинною структурою, побудованою з рибонуклеїнових кислот (рибосомних РНК) і білків. Для того щоб синтезувати білок, рибосома має бути забезпечена:
1. Програмою, що задає порядок чергування амінокислотних залишків поліпептидної ланцюга білка.
2. Амінокислотним матеріалом, з якого слід будувати білок.
3. Енергією.
Сама рибосома має каталітичну (ензиматичну) функцію, відповідальну за утворення пептидних зв'язків і, відповідно, полімеризацію амінокислотних залишків у поліпептидний ланцюг білка.
Програма, що задає порядок чергування амінокислотних залишків в поліпептидному ланцюзі білка, виходить від дезоксирибонуклеїнової кислоти (ДНК), т. Е. З генома клітини. Синтезовані ланцюги РНК компліментарні одному з ланцюгів ДНК і, таким чином, точно відтворюють дезоксирибонуклеотидну послідовність іншого ланцюга ДНК у своїй рибонуклеотидній послідовності. Процес такого копіювання гена, здійснюваний ферментом РНК-полімеразою, отримав назву транскрипції. РНК протягом синтезу та після нього, особливо в еукаріотичних клітинах, може піддаватися ряду додаткових змін, званих процесингом, в ході яких з неї можуть бути вирізані певні шматки нуклеотидної послідовності. РНК, що виходить, надходить далі в рибосоми в якості програми, що визначає амінокислотну послідовність в синтезованому білку. Вона називається інформаційною або "месенджером" РНК (мРНК). Таким чином, саме транскрипція генів та утворення мРНК забезпечують потік інформації від ДНК до рибосом.
Початковим матеріалом, з якого будується білок, є амінокислоти. Однак вільні амінокислоти не використовуються рибосомою, Для того щоб служити субстратом для рибосоми, амінокислота повинна бути активована за участю сполученого розщеплення АТФ і акцептована (ковалентно приєднана) спеціальною молекулою РНК, званої трансферної або транспортної РНК (тРНК), за допомогою ферменту синтетези. Отримані аміноацил-тРНК надходять у рибосому як субстрат для синтезу білка. Крім того, енергія хімічного зв'язку між амінокислотним залишком та тРНК використовується для реакції утворення пептидного зв'язку в рибосомі. Таким чином, активація амінокислот та утворення аміноацил-тРНК забезпечують потік як матеріалу, так і енергії для рибосомного синтезу білка.
Ці три потоки (інформації, матеріалу та енергії) зустрічаються в рибосомі. Сприймаючи їх, рибосома здійснює переклад, або трансляцію, генетичної інформації з нуклеотидної мови послідовності мРНК на мову амінокислотної послідовності синтезованого поліпептидного ланцюга білка. Якщо уявити це в молекулярних термінах, то рибосома послідовно сканує ланцюг мРНК (рухається вздовж неї) і теж послідовно вибирає з середовища аміноацил-тРНК, в результаті чого специфічність аміноацильного залишку вибирається рибосомою аміноацил-тРНК щоразу детермінується специфічністю комбінації. відрізка мРНК. Отже, виникає проблема генетичного коду: які комбінації нуклеотидів детермінують, т. е. чи кодують кожну з 20 амінокислот, з яких будуються молекули білків?
Рух рибосоми вздовж ланцюга мРНК (або, іншими словами, пропускання ланцюга мРНК крізь рибосому) задає строгий тимчасовий порядок входження в рибосому різних аміноацил-тРНК відповідно до порядку розташування нуклеотидних комбінацій, що кодують, уздовж мРНК. Аміноацильний залишок обраного аміноацил-тРНК щоразу ковалентно приєднується рибосомою до зростаючого поліпептидного ланцюга. Деацильована тРНК звільняється з рибосоми у розчин. Так послідовно, крок за кроком, будується поліпептидний ланцюг білка (див. схему 1).
Біосинтез білка.
Пластичний обмін (асиміляція чи анаболізм) – сукупність реакцій біологічного синтезу. Назва цього виду обміну відбиває його сутність: з речовин, які у клітину з поза, утворюються речовини, подібні речовинам клітини.
Розглянемо одну з найважливіших форм пластичного обміну – біосинтез білків. Біосинтез білківздійснюється у всіх клітинах про -і еукаріотів. Інформація про первинну структуру (порядок амінокислот) білкової молекули закодована послідовністю нуклеотидів у ділянці молекули ДНК - гені.
Ген-це ділянка молекули ДНК, що визначає порядок амінокислот в молекулі білка. Отже, від нуклеотидів у гені залежить порядок амінокислот в поліпептиді, тобто. його первинна структура, від якої у свою чергу залежать всі інші структури, властивості та функції білкової молекули.
Система запису генетичної інформації в ДНК (і РНК) у вигляді певної послідовності нуклеотидів називається генетичним кодом. Тобто. одиниця генетичного коду (кодон) - це триплет нуклеотидів у ДНК або РНК, що кодує одну амінокислоту.
Усього генетичний код включає 64 кодони, з них 61 кодуючий і 3 некодуючі (кодони-термінатори, що свідчать про закінчення процесу трансляції).
Кодони-термінатори і - РНК: УАА, УАГ, УГА, в ДНК: АТТ, АТЦ, АЦТ.
Початок процесу трансляції визначає кодон-ініціатор (АУГ, ДНК - ТАЦ), що кодує амінокислоту метіонін. Цей кодон першим входить у рибосому. Згодом метіонін, якщо він не передбачений як перша амінокислота даного білка, відщеплюється.
Генетичний код має характерні властивості.
1. Універсальність – код однаковий для всіх організмів. Один і той же триплет (кодон) в будь-якому організмі кодує ту саму амінокислоту.
2. Специфічність – кожен кодон шифрує лише одну амінокислоту.
3. Виродженість – більшість амінокислот можуть кодуватися кількома кодонами. Виняток становлять 2 амінокислоти - метіонін і триптофан, що мають лише за одним варіантом кодону.
4. Між генами є «розділові знаки» - три спеціальні триплети (УАА, УАГ, УГА), кожен з яких позначає припинення синтезу поліпептидного ланцюга.
5. Усередині гена «розділових знаків» немає.
Для того, щоб синтезувався білок, інформація про послідовність нуклеотидів у його первинній структурі має бути доставлена до рибосом. Цей процес включає два етапи – транскрипцію та трансляцію.
Транскрипція(переписування) інформації відбувається шляхом синтезу на одному з ланцюгів молекули ДНК одноланцюжкової молекули РНК, послідовність нуклеотидів якої точно відповідає послідовності нуклеотидів матриці – полінуклеотидного ланцюга ДНК.
Вона (і - РНК) є посередником, що передає інформацію від ДНК до місця збирання молекул білка в рибосомі. Синтез і РНК (транскрипція) відбувається наступним чином. Фермент (РНК - полімераза) розщеплює подвійний ланцюжок ДНК, і однією з її ланцюгів (кодирующей) за принципом комплементарності вибудовуються нуклеотиди РНК. Синтезована таким чином (матричний синтез) молекула і РНК виходить в цитоплазму, і на один її кінець нанизуються малі субодиниці рибосом.
Другий етап у біосинтезі білка - трансляція- це переклад послідовності нуклеотидів у молекулі та - РНК у послідовність амінокислот у поліпептиді. У прокаріотів, що не мають оформленого ядра, рибосоми можуть зв'язуватися з новоствореною молекулою і - РНК відразу ж після її відділення від ДНК або навіть до повного завершення її синтезу. У еукаріотів і - РНК спочатку має бути доставлена через ядерну оболонку в цитоплазму. Перенесення здійснюється спеціальними білками, які утворюють комплекс з молекулою і РНК. Крім функцій перенесення ці білки захищають і - РНК від дії, що пошкоджує цитоплазматичних ферментів.
У цитоплазмі однією з кінців і - РНК (а саме на той, з якого починається синтез молекули в ядрі) вступає рибосома і починається синтез поліпептиду. У міру просування молекулою РНК рибосома транслює триплет за триплетом, послідовно приєднуючи амінокислоти до зростаючого кінця поліпептидного ланцюга. Точна відповідність амінокислоти коду триплету і РНК забезпечується т РНК.
Транспортні РНК (т – РНК) «приносять» амінокислоти у велику субодиницю рибосоми. Молекула т – РНК має складну конфігурацію. На деяких ділянках її між комплементарними нуклеотидами утворюються водневі зв'язки, і молекула формою нагадує лист конюшини. На її верхівці розташований триплет вільних нуклеотидів (антикодон), який відповідає певній амінокислоті, а основа є місцем прикріплення цієї амінокислоти (рис. 1).
Мал. 1. Схема будови транспортної РНК: 1 – водневі зв'язки; 2 – антикодон; 3-місце прикріплення амінокислоти.
Кожна т – РНК може переносити лише свою амінокислоту. Т-РНК активується спеціальними ферментами, приєднує свою амінокислоту та транспортує її в рибосому. Усередині рибосоми в кожний момент знаходиться всього два кодони і-РНК. Якщо антикодон т-РНК є комплементарним кодону і-РНК, відбувається тимчасове приєднання т-РНК з амінокислотою до і-РНК. До другого кодону приєднується друга т-РНК, яка несе свою амінокислоту. Амінокислоти розташовуються поруч великої субодиниці рибосоми, і з допомогою ферментів з-поміж них встановлюється пептидна зв'язок. Одночасно руйнується зв'язок між першою амінокислотою та її т-РНК, і т-РНК йде з рибосоми за наступною амінокислотою. Рибосома переміщається однією триплет, і процес повторюється. Так поступово нарощується молекула поліпептиду, в якій амінокислоти розташовуються в суворій відповідності з порядком триплетів, що кодують їх (матричний синтез) (рис. 2).

Мал. 2. Схема бісинтезу білка: 1 – і-РНК; 2 - субодиниці рибосоми; 3 – т-РНК з амінокислотами; 4 – т-РНК без амінокислот; 5 – поліпептид; 6 - кодон та-РНК; 7-антикодон т-РНК.
Одна рибосома здатна синтезувати повний поліпептидний ланцюг. Проте, нерідко однією молекулою і-РНК рухається кілька рибосом. Такі комплекси називаються полірибосомами. Після завершення синтезу поліпептидний ланцюжок відокремлюється від матриці – молекули і-РНК, згортається в спіраль і набуває властивої їй (вторинної, третинної або четвертинної) структури. Рибосоми працюють дуже ефективно: протягом 1с бактеріальна рибосома утворює поліпептидний ланцюг із 20 амінокислот.
Біосинтез білків (поліпептидів) є надзвичайно складним та дивовижним процесом. Біосинтез білків активно протікає у всіх органах та тканинах, виключаючи еритроцити. Багато клітин синтезують білки на «експорт» (клітини печінки, підшлункової залози), і в цьому випадку вони містять дуже велику кількість рибосом. У тваринній клітині число рибосом досягає 10 5 діаметр рибосоми дорівнює 20 нм.
Процес синтезу білка відбувається всередині клітин на поверхні рибосом, які є комплексами з двох субодиниць з константою седиментації 60S і 40S, що функціонують як єдине ціле. У рибосомі білок становить 30-35% та рибосомальна РНК – 65-70%. У рибосомі розрізняють аміноацильні та пептидильні ділянки. Перший служить для фіксації комплексу активної амінокислоти і тРНК, що надходить на рибосому, а другий фіксує поліпептидний ланцюг, пов'язаний з іншою тРНК. Субодиниці рибосом синтезуються в ядерці ядра на матриці ДНК.
Сутність процесу синтезу білка представляє схема:
Білоксинтезуюча система включає рибосоми, нуклеїнові кислоти, набір з 20 амінокислот, різні ферменти, АТФ, ГТФ, іони магнію, близько 200 різних некаталітичних білкових факторів.
Молекула білка - довгий ланцюг амінокислотних залишків, що налічує в середньому від 100 до 500 амінокислот. Програма синтезу кожного білка зберігається у молекулі дезоксирибонуклеїнової кислоти (ДНК). Молекула ДНК - полімер, мономерами якого є нуклеотиди. Послідовність азотистих основ у молекулі ДНК визначає послідовність амінокислот у молекулі білка.
У молекулі ДНК є чотири види азотистих основ: аденін (А), гуанін (Г), цитозин (Ц) та тимін (Т). Послідовність із трьох підстав (триплет) становить кодон, якому відповідає одна певна амінокислота.
Нуклеїнові кислоти – ДНК та РНК – обов'язкові компоненти біосинтезу білків. ДНК відповідає за збереження генетичної інформації, тоді як РНК визначає передачу цієї інформації та реалізацію у вигляді молекул білка. Можна стверджувати, що головна функція ДНК – це збереження генотипу, а РНК – вираз цього генотипу.
У кількісному плані у клітині переважає рибосомальна РНК (рРНК). рРНК має спіралізовані ділянки, містить модифіковані нуклеотиди (наприклад, 2-метил-рибозу). рРНК становить близько 80% від загальної кількості РНК у клітині. Другий вид РНК у клітині представлений транспортною РНК (тРНК), яка, як та інші види РНК, синтезується в ядрі. На її частку припадає 10-15% загальної кількості РНК у клітині. Виявлено понад 60 різних тРНК. Тож транспорту окремих амінокислот існує кілька різних тРНК. Для кожної амінокислоти у клітині є принаймні одна специфічна тРНК. Молекули тРНК порівняно дрібні. У їх структурі 75-93 рибонуклеїдів.
Амінокислота приєднується до вільної 3-ОН-групи кінцевого мононуклеотиду тРНК, представленої завжди аденіловою кислотою. тРНК має й іншу важливу ділянку - антикодон, за допомогою якого комплекс амінокислоти та тРНК дізнається певну послідовність із трьох нуклеотидів у матричній РНК (кодон). Антикодон та кодон комплементарно з'єднуються за допомогою водневих зв'язків.
Якщо носієм спадкової інформації в клітині є ДНК, яка зосереджена в ядрі, але синтез білка відбувається в цитоплазмі, то, отже, має бути певний посередник, який передає цю інформацію цитоплазму клітини. Цим посередником виявилася інформаційна чи матрична РНК (мРНК). Перед мРНК припадає 2% загальної кількості РНК клітини. Молекули мРНК найдовші (включають до 5 тис. Нуклеотидів). мРНК також містить чотири види азотистих основ. З них три (А, Г, Ц) такі ж, як у ДНК, а четверте – урацил.
Інформація, закодована в мРНК, необхідна синтезу молекули білка, що відбувається на рибосомах. Синтез мРНК у ядрі клітини дуже швидкий, що необхідне активного біосинтезу білкових молекул. мРНК утворюється на одній із ниток ДНК ядра. При цьому двоспіральна структура ДНК розкручується і за участю ДНК-залежної РНК-полімерази за принципом комплементарності відбувається синтез мРНК:

Схема синтезу мРНК
Принцип комплементарності означає, що аденіну на спіралі ДНК відповідає урацил мРНК, тиміну – аденін, а гуаніну – цитозин. Отже, мРНК зчитує інформацію із ДНК.
Стадія ДНК - РНК, таким чином, визначає синтез молекули мРНК, в якій нуклеотидна послідовність комплементарна певному ділянці (гену) ДНК. Цей процес зветься транскрипції. Потім мРНК надходить на рибосому, поєднуючись з її субодиницями. Одна молекула мРНК фіксується на множині рибосом одночасно, утворюючи так звані полісоми. Наявність полісом підвищує ефективність та швидкість використання мРНК.
Синтез поліпептидного ланцюга певного складу відбувається на матриці мРНК. Процес передачі з мРНК на білок отримав назву трансляції. Стадія «РНК -> білок» є процесом синтезу білка, що направляється мРНК. Таким чином, передача інформації завжди йде в напрямку ДНК - РНК - білок.
Процес трансляції включає такі етапи:
Цей же фермент бере участь у фіксації попередньо активованої амінокислоти в положення 2 або 3 рибози останнього нуклеотиду тРНК:
У вигляді даного комплексу амінокислота транспортується рибосому, де відбувається синтез білкової молекули. Аміноацил-тРНК-синтетаза специфічна, вона здатна впізнавати як амінокислоту, і тРНК. У клітині, таким чином, є не менше 20 різних синтетаз відповідно до числа а-амінокислот.
2. тРНК, пов'язана ефірним зв'язком з певною амінокислотою, надходить на рибосому і взаємодіє з мРНК за типом комплементарності між специфічним триплетом нуклеотидів мРНК, названим кодоном, і їй комплементарним специфічним триплетом нуклеотидів (антикодоном) тР. Таким чином, кожен кодон мРНК відповідає специфічній фіксації однієї амінокислоти в пептидному ланцюзі за допомогою антикодону тРНК. Рибосома пересувається вздовж молекули мРНК, послідовно зчитуючи всі кодони, встановлюючи таким чином порядок розташування всіх амінокислот, що доставляють до місця синтезу.
Синтез молекули білка йде у напрямку від вільної аміногрупи до вільної карбоксильної групи амінокислоти. Зазвичай початковою амінокислотою в синтезі поліпептидного ланцюга є метіонін, для якого кодоном є нуклеотидна послідовність АУГ мРНК.
Ініціація синтезу поліпептиду починається при фіксації двох антикодонів тРНК за відповідними кодонами мРНК. Процес вимагає наявності джерела енергії, яким служить ГТФ, а також участі цілого ряду білкових факторів ініціації та пептиділтрансферази.
За участю даного ферменту швидкість освіти ковалентних зв'язківдосягає 1200 амінокислот/хв/рибосом.

Схема ініціації синтезу поліпептиду
3. Після утворення дипептиду «ненавантажена» тРНК залишає рибосому і здатна доставляти нові молекули амінокислот, а мРНК просувається щодо рибосоми (полісоми) на три нуклеотиди. В результаті переміщення (транслокації) вільний кодон займає положення для впізнавання чергової молекули тРНК. Отже, у стадії елонгації відбувається послідовне приєднання по одній амінокислоті до поліпептидного ланцюга у суворій відповідності до порядку кодонів молекули мРНК.
Поліпептидний ланцюг, що подовжується, з однією молекулою тРНК фіксується з великою субодиницею рибосоми. Приєднання кожної додаткової амінокислоти до поліпептидного ланцюга відбувається за рахунок взаємозв'язку аміногрупи амінокислоти, що приєднується в комплексі з тРНК і карбоксильної групи пептиду.
4. Термінація або завершення синтезу поліпептидної молекули залучає певні кодони термінації «без сенсу» та білкові фактори термінації. Відомі три кодони (УАГ, УГА, УАА), які не кодують, не пов'язують якусь амінокислоту, так як в клітині не існує антикодонів тРНК, комплементарних до них. Теоретично лише один кодон "без сенсу", відомий полісомою під час проходження в напрямку 5-3 мРНК, повинен зупинити синтез молекули білка.
Наявність термінуючого кодону у будь-якій ділянці мРНК означає закінчення білкового синтезу. В результаті полісому розпадається, невикористана мРНК гідролізується полінукле- леотидфосфорілазою, а субодиниці рибосом готуються до початку синтезу нової молекули білка.
мРНК може неодноразово брати участь у процесі біосинтезу білка. Тривалість функціонування молекули мРНК неоднакова у різних організмів. Вона може коливатися від кількох хвилин до кількох діб.
5. У ДНК закодовано лише первинну структуру білка. Тому синтезовані на рибосомах молекули білків ще мають остаточно завершеного стану. Вони представляють первинні поліпептиди, які потім зазнають численних модифікацій (асоціації мономерів з утворенням олігомерів, приєднання коферментів, хімічні перетворення), що змінюють структуру білків і, отже, їх активність.
Вторинна та третинна структури не кодовані, вони визначаються властивостями первинної структури, а це означає, що та чи інша форма білкової молекули залежить від послідовності амінокислот та можливостей їх взаємодії між собою. Структурні модифікації білків, що синтезуються, мають місце ще на рівні рибосом або після завершення синтезу в результаті приєднання різних функціональних груп.
Розглянута схема передачі у вигляді
може в окремих випадках змінюватись. Так, у вірусів, які не містять ДНК, інформація закладена в РНК. При проникненні вірусу клітину ця інформація передається на ДНК клітини, а остання вже синтезує мРНК, на матриці якої синтезуються вірусні білки. Такий процес носить назву зворотної транскрипції, і схема передачі в цьому випадку буде наступною:
Поки зберігається послідовність нуклеотидів ДНК і, отже, мРНК, характер білка, що знову синтезується, залишається незмінним.
Необхідна генетична інформація для синтезу білка може бути представлена аналогічно до запису людської мови, яка складається з послідовності букв, що формують слова та речення. У генетичній мові, однак, є лише чотири літери - чотири основи (аденін, гуанін, урацил, цитозин).
Генетичний код включає трилітерні слова. Чотири підстави в даному випадку (43) дають 64 варіанти (слова), яких більш ніж достатньо, щоб кодувати 20 амінокислот. Таким чином, 64 кодони і складають генетичний код (табл. 3).
Аналіз генетичного коду показує, що з різних амінокислот є різне число кодонов. Наприклад, метіонін і триптофан мають лише один кодон, тоді як аргінін, лейцин, серин мають по шість кодонів. Наявність кількох кодонів для однієї амінокислоти відображає «виродженість» коду. Отже, та сама амінокислота може кодуватися кількома за своєю будовою нуклеотидними триплетами. У той же час кожному триплету відповідає цілком певна амінокислота в поліпептидному ланцюгу, що синтезується.
Таблиця 3
Генетичний код
|
нуклеотид |
Другий нуклеотид |
нуклеотид |
|||
Генетичний код універсальний і однаковий у видів різного рівня розвитку (людина, тварини, рослини, мікроорганізми). Універсальність коду свідчить, що всі живі організми минулого мали єдиного предка.
Окремі амінокислоти (оксипролін, оксилизин), наприклад, не мають кодону і утворюються за допомогою хімічних реакційвже після синтезу поліпептидного ланцюга. Цей процес отримав назву посттрансляційної модифікації та дуже важливий для правильного функціонування кожного білка.
Безглузді кодони (УАА, УАГ, УГА) не кодують амінокислоти, проте реально є сигналом закінчення синтезу білкової молекули.
Таким чином, мРНК є безпосереднім переносником генетичної інформації з ядра на рибосому цитоплазми. Одна рибосома займає на мРНК ділянку довжиною близько 80 нуклеотидів і здатна каталізувати приблизно 100 пептидних зв'язків за хвилину (Северін Е. С. та ін., 2011).
Синтезовані білкові молекули можуть піддаватися структурним модифікаціям на рівні рибосом або після завершення синтезу в результаті приєднання різних функціональних груп. У цитоплазмі мРНК має порівняно короткий період. Деяка кількість мРНК синтезується та запасається у неактивній формі, будучи готовою для швидкого синтезу білка. Оскільки інформація мРНК пов'язана з лінійною послідовністю нуклеотидів, цілісність цієї послідовності є надзвичайно важливою. Будь-яка втрата або зміна порядку нуклеотидів може змінити синтез білка. На сьогодні встановлено цілу низку інгібіторів реплікації ДНК у клітинах організму (антибіотики, хімічні отрути, антивірусні препарати). Пошкодження в послідовності пуринових або піримідинових основ у гені отримали назву мутації.
Заміна лише одного нуклеотиду в кодоні (мутація) призводить до зміни кодування однієї амінокислоти на іншу. Наприклад, мутація, пов'язана із заміною глутамінової кислоти на валін у молекулі гемоглобіну, призводить до синтезу гемоглобіну, що викликає серпоподібну анемію. Сьогодні відомо понад 200 мутацій поліпептидного кола молекули гемоглобіну людини. Часто мутагенами є речовини (нітрозаміни, наприклад), що змінюють структуру азотистих основ, що призводить до зміни характеру комплементарності основ. Ультрафіолетове опромінення викликає конденсацію залишків тиміну з утворенням тімінових димерів. На щастя, від шкідливого впливу ультрафіолетових променів тварини захищені шаром озону атмосфери.
Багато антибіотиків, що використовуються у ветеринарній практиці, інгібують бактеріальний синтез білка (лінкоміцин, еритроміцин, хлорамфенікол) ще на стадії трансляції. При цьому мікробна клітина гине чи зупиняє свій розвиток. Такі антибіотики, як тетрацикліни, не впливають на рибосомальний синтез у клітинах вищих тварин. Пеніциліни не є прямими інгібіторами синтезу білка, проте їх ефекти пригнічення бактерій пов'язані з блокуванням синтезу гексапептидів клітинної стінки. Слід зазначити, що синтез білка відбувається не тільки на рибосомах, а й у мітохондріях. Мітохондрії мають повний та незалежний апарат синтезу білка для своїх потреб, хоча не всі мітохондріальні білки синтезуються в цих органелах. РНК мітохондрій становлять лише 3% від кількості РНК клітини. Рибосоми мітохондрій менші за розмірами, ніж цитоплазматичні. Кодон УГА як термінатор синтезу білка в цитоплазмі використовується в мітохондріях поряд з кодоном УГГ для кодування амінокислоти.
Синтезовані на рибосомах білки не мають остаточно завершеного стану. Вони представляють первинні поліпептиди, які потім зазнають численних модифікацій (асоціації мономерів з утворенням олігомерів, приєднання коферментів, хімічні перетворення), що модифікують структуру білка і, отже, його активність.
Картинка 9 із презентації «Біосинтез білка»до уроків біології на тему «Біосинтез білка»
Розміри: 960 х 720 пікселів, формат: jpg. Щоб безкоштовно скачати картинку для уроку біології, клацніть на зображенні правою кнопкою миші і натисніть «Зберегти зображення як...». Для показу картинок на уроці Ви можете також безкоштовно скачати презентацію «Біосинтез білка.pptx» повністю з усіма картинками в zip-архіві. Розмір архіву – 1719 КБ.
Завантажити презентацію«Функції білків» - Так відбувається прийом сигналів із зовнішнього середовища та передача інформації в клітину. При розпаді 1 г білка до кінцевих продуктів виділяється 17,6 кДж. Що таке ренатурація? Підіб'ємо підсумки: 9. Каталітична. Процес відновлення структури білка після денатурації називається ренатурацією. Піменов А.В. Білки є одним із джерел енергії в клітині.
«Білки речовина» - Наприклад: колаген. Викладач біології: Болдирєва Л. А. Відомо 20 АК, з яких будуються білки. . Приклад: варене яйце. Амінокислота – органічна речовина, Нерозчинні білки – фібрилярні. Харчові білки. . Захисні білки. Структура білка. Використовуються організмом для руху. Енергетичні білки.
«Білки та його функції» - Каталітична роль. Двигуна функція. Концепція білки. Гідроліз білків зводиться до розщеплення поліпептидних зв'язків: Висновок: З білків побудовано кровоносні судини, сухожилки, волосся. Будова та функції білка. Хімічні властивостібілків. білки беруть участь в утворенні оболонки клітини, органоїдів та мембран клітини.
"Біосинтез білка" - Список літератури. Вступ. 4. Зміст. Біосинтез білків у живій клітині. 7. 10. 9. Схема рослинної та тваринної клітин. 5. 6. 1. 8. 2. 3.
"Біосинтез білків" - Трансляція (лат. Перенесення, переклад). Транскрипція (лат. переписування). Перевір себе. Значення білків. Зміст. Енергетика біосинтезу. Роль ферментів. Синтез поліпептидного ланцюга на рибосомі. 5. Яка послідовність нуклеотидів і-РНК, записаної на відрізку ДНК: Т-А-Ц-Г-Г-А-Т-Ц-А-Ц-Г-А -Г-Т-Г-Ц-Т А-У-Г-Ц-Г-У-А-Г-У-Г-Ц-У А-У-Г-Ц-Ц-У-А-Г-У -Г-Ц-У.
"Біосинтез білка біологія" - Микола Костянтинович Кольцов (1872-1940). А. Г. Основною функцією рибосом є синтез білків. Центральна догма (основний постулат) молекулярної біології – матричний синтез. Ц. Антикодон - триплет нуклеотидів на верхівці тРНК. Біосинтез білка. Після завершення синтезу іРНК розпадається на нуклеотиди.
Всього у темі 8 презентацій
© А.С. Спірін
БІОСИНТЕЗ БІЛКІВ, СВІТ РНКА.С. Спірін
Спірін Олександр Сергійович– академік, директор Інституту білка РАН, член Президії РАН.
Майже півстоліття тому, 1953 р., Д. Вотсон і Ф. Крик відкрили принцип структурної (молекулярної) організації генної речовини - дезоксирибонуклеїнової кислоти (ДНК). Структура ДНК дала ключ до механізму точного відтворення – редуплікації – генної речовини. Так виникла нова наука – молекулярна біологія. Було сформульовано так звану центральну догму молекулярної біології: ДНК Ю РНК Ю білок. Сенс її полягає в тому, що генетична інформація, записана в ДНК, реалізується у вигляді білків, але не безпосередньо, а за допомогою спорідненого полімеру - рибонуклеїнову кислоту (РНК), і цей шлях від нуклеїнових кислот до білків необоротний. Таким чином ДНК синтезується на ДНК, забезпечуючи власну редуплікацію, тобто відтворення вихідного генетичного матеріалу в поколіннях; РНК синтезується на ДНК, у результаті відбувається переписування, чи транскрипція, генетичної інформації у форму численних копій РНК; молекули РНК є матрицями для синтезу білків - генетична інформація транслюється у форму поліпептидних ланцюгів. У спеціальних випадках РНК може переписуватись у форму ДНК ("зворотна транскрипція"), а також копіюватися у вигляді РНК (реплікація), але білок ніколи не може бути матрицею для нуклеїнових кислот (докладніше див.).
Отже, саме ДНК визначає спадковість організмів, тобто набір білків, що відтворюється в поколіннях, і пов'язаних з ними ознак. Біосинтез білка є центральним процесом живої матерії, а нуклеїнові кислоти забезпечують його, з одного боку, програмою, що визначає весь набір і специфіку білків, що синтезуються, а з іншого - механізмом точного відтворення цієї програми в поколіннях. Отже, походження життя у її сучасній клітинній формі зводиться до виникнення механізму успадкованого біосинтезу білків.
БІОСИНТЕЗ БІЛКІВ
Центральна догма молекулярної біології постулює лише шлях передачі генетичної інформації від нуклеїнових кислот до білків і, отже, до властивостей та ознак живого організму. Вивчення механізмів реалізації цього шляху протягом десятиліть, що послідували за формулюванням центральної догми, розкрило набагато різноманітніші функції РНК, ніж бути лише переносником інформації від генів (ДНК) до білків і служити матрицею для синтезу білків.
На рис. 1 представлена загальна схема біосинтезу білка у клітині. РНК-посередник(messenger RNA, матрична РНК, мРНК), що кодує білки, про яку йшлося вище, - це лише один із трьох головних класів клітинних РНК. Основну їхню масу (близько 80%) становить інший клас РНК - рибосомні РНК,які утворюють структурний каркас та функціональні центри універсальних білок-синтезуючих частинок – рибосом. Саме рибосомні РНК відповідальні - як у структурному, і у функціональному відношенні - формування ультрамікроскопічних молекулярних машин, званих рибосомами. Рибосоми сприймають генетичну інформацію як молекул мРНК і, будучи запрограмовані останніми, роблять білки у точній відповідності з цією програмою.
Проте, щоб синтезувати білки, лише інформації чи програми недостатньо - потрібен ще й матеріал, з якого їх можна робити. Потік матеріалу для синтезу білків йде в рибосоми за допомогою третього класу клітинних РНК. РНК-переносників(Трансфер RNA, транспортні РНК, тРНК). Вони ковалентно пов'язують - акцептують - амінокислоти, які є будівельним матеріалом для білків, і у вигляді аміноацил-тРНК надходять у рибосоми. У рибосомах аміноацил-тРНК взаємодіють з кодонами - тринуклеотидними комбінаціями - мРНК, внаслідок чого відбувається декодування кодонів у процесі трансляції.
Рибонуклеїнові кислоти
Отже, маємо набір основних клітинних РНК, визначальних основний процес сучасної живої матерії - біосинтез білка. Це мРНК, рибосомні РНК та тРНК. РНК синтезуються на ДНК за допомогою ферментів - РНК-полімераз, що здійснюють транскрипцію - переписування певних ділянок (лінійних відрізків) двотяжової ДНК у форму однотяжової РНК. Ділянки ДНК, що кодують клітинні білки, переписуються у вигляді мРНК, тоді як для синтезу численних копій рибосомної РНК і тРНК є спеціальні ділянки клітинного геному, з яких йде інтенсивне переписування без подальшої трансляції білки.
Хімічна структура РНК.
Хімічно РНК дуже схожа на ДНК. Обидві речовини – це лінійні полімери нуклеотидів. Кожен мономер - нуклеотид - являє собою фосфорильований N-глікозид, побудований з залишку п'ятивуглецевого цукру - пентози, що несе фосфатну групу на гідроксильній групі п'ятого вуглецевого атома (складноефірний зв'язок) і азотисту основу при першому вуглецевому атомі (N-глікозину) Головне хімічне різницю між ДНК і РНК у тому, що цукровий залишок мономеру РНК - це рибоза, а мономеру ДНК - дезоксирибоза, що є похідним рибози, у якому відсутня гідроксильна група при другому вуглецевому атомі (рис. 2).
 Мал. 2.Хімічні формули залишків одного з рибонуклеотидів - уридилової кислоти (U) та гомологічного йому дезоксирибонуклеотиду - тимідилової кислоти (dT) |
Азотистих основ і в ДНК, і в РНК чотири види: два пуринових - аденін (А) і гуанін (G) -і два піримідинових - цитозин (С) і урацил (U) або його метильоване похідне тімін (Т). Урацил характерний для мономерів РНК, а тімін - для мономерів ДНК, і це друга відмінність РНК та ДНК. Мономери – рибонуклеотиди РНК або дезоксирибонуклеотиди ДНК – утворюють полімерний ланцюг за допомогою формування фосфодіефірних містків між цукровими залишками (між п'ятим та третім атомами вуглецю пентози). Таким чином, полімерний ланцюг нуклеїнової кислоти - ДНК або РНК - може бути представлений як лінійний сахаро-фосфатний кістяк з азотистими основами як бічні групи. Макромолекулярна структура РНК. Принципова макроструктурна відмінність двох типів нуклеїнових кислот полягає в тому, що ДНК - це єдина подвійна спіраль, тобто макромолекула з двох пов'язаних комплементарно полімерних тяжів, спірально закручених навколо загальної осі (див. [ , ]), а РНК - однотяжовий полімер. У той же час взаємодії бічних груп - азотистих основ - один з одним, а також з фосфатами і гідроксилами сахаро-фосфатного кістяка призводять до того, що однотяжовий полімер РНК згортається на себе і скручується в компактну структуру, подібно до згортання поліпептидного ланцюга білка в компактну глобулу . Таким чином, унікальні нуклеотидні послідовності РНК можуть формувати унікальні просторові структури. |
Вперше специфічна просторова структура РНК була продемонстрована при розшифровці атомної структури однієї з тРНК у 1974 р. [ , ] (Рис. 3). Згортання полімерного ланцюга тРНК, що складається з 76 нуклеотидних мономерів, призводить до формування дуже компактного глобулярного ядра, з якого під прямим кутом стирчать два виступи. Вони є короткі подвійні спіралі на кшталт ДНК, але організовані з допомогою взаємодії ділянок однієї й тієї ж ланцюга РНК. Один із виступів є акцептором амінокислоти і бере участь у синтезі поліпептидного ланцюга білка на рибосомі, а інший призначений для комплементарної взаємодії з триплетом (кодоном) мРНК, що кодує, в тій же рибосомі. Тільки така структура здатна специфічно взаємодіяти з білком-ферментом, що навішує амінокислоту на тРНК, і рибосомою в процесі трансляції, тобто специфічно "впізнаватись" ними.

Мал. 3.Атомна (ліворуч) та скелетна (праворуч) моделі фенілаланінової тРНК дріжджів
Вивчення ізольованих рибосомних РНК дало наступний разючий приклад формування компактних специфічних структур ще більш довгих лінійних полімерів цього типу. Рибосома складається з двох нерівних частин - великої та малої рибосомних субчастинок (субодиниць). Кожна субчастиця побудована з однієї високополімерної РНК та цілого ряду різноманітних рибосомних білків. Довжина ланцюгів рибосомних РНК дуже значна: так, РНК малої субчастинки бактеріальної рибосоми містить понад 1500 нуклеотидів, а РНК великої субчастинки - близько 3000 нуклеотидів. У ссавців, включаючи людину, ці РНК ще більше - близько 1900 нуклеотидів і понад 5000 нуклеотидів у малій та великій субчастинках відповідно.
Було показано, що ізольовані рибосомні РНК, відокремлені від своїх білкових партнерів та отримані в чистому вигляді, самі здатні спонтанно згортатися в компактні структури, за своїми розмірами та формою схожі на рибосомні субчастинки]. Форма великої та малої субчастиць різна, і відповідно відрізняється форма великої та малої рибосомних РНК (рис. 4). Таким чином, лінійні ланцюги рибосомної РНК самоорганізуються в специфічні просторові структури, що визначають розміри, форму і, мабуть, внутрішній пристрій рибосомних субчастинок, а отже, і всієї рибосоми.
Мінорні РНК. У міру вивчення компонентів живої клітини та окремих фракцій тотальної клітинної РНК з'ясовувалося, що трьома головними видами РНК справа не обмежується. Виявилося, що у природі існує безліч інших видів РНК. Це насамперед так звані "малі РНК", які містять до 300 нуклеотидів, часто з невідомими функціями. Як правило, вони асоційовані з одним або декількома білками і представлені в клітці у вигляді рибонуклеопротеїдів - "малих РНП".
Малі РНК присутні у всіх відділах клітини, включаючи цитоплазму, ядро, ядерце, мітохондрії. Більшість тих малих РНП, функції яких відомі, бере участь у механізмах посттранскрипційної обробки основних видів РНК (RNA processing) - перетворенні попередників мРНК на зрілі мРНК (сплайсинг), редагуванні мРНК, біогенезі тРНК, дозріванні рибосомних РНК. Один з найбільш багато представлених у клітинах видів малих РНП (SRP) відіграє ключову роль у транспорті білків, що синтезуються, через клітинну мембрану. Відомі види малих РНК, що виконують регуляторні функції у трансляції. Спеціальна мала РНК входить до складу найважливішого ферменту, відповідального за підтримку редуплікації ДНК у поколіннях клітин – теломерази. Слід сказати, що їх молекулярні розміри можна порівняти з розмірами клітинних глобулярних білків. Таким чином, поступово стає ясно, що функціонування живої клітини визначається не тільки різноманіттям білків, що синтезуються в ній, але і присутністю багатого набору різноманітних РНК, з яких малі РНК значною мірою імітують компактність і розміри білків.
Рибозими. Все активне життя побудована на обміні речовин - метаболізмі, і всі біохімічні реакції метаболізму відбуваються з належними для забезпечення життя швидкостями тільки завдяки високоефективним специфічним каталізаторам, створеним еволюцією. Упродовж багатьох десятиліть біохіміки були впевнені, що біологічний каталіз завжди і всюди здійснюється білками, які називаються ферментами, або ензимами.І ось у 1982-1983 pp. було показано, що в природі є види РНК, які, подібно до білків, мають високоспецифічну каталітичну активність [ , ]. Такі РНК-каталізатори було названо рибозимами.Уявленню про винятковість білків у каталізі біохімічних реакцій настав кінець.
В даний час рибосому також прийнято розглядати як рибозим. Справді, всі наявні експериментальні дані свідчать, що синтез поліпептидної ланцюга білка в рибосомі каталізується рибосомної РНК, а чи не рибосомними білками. Ідентифіковано каталітичну ділянку великої рибосомної РНК, відповідальну за каталіз реакції транспептидації, за допомогою якої здійснюється нарощування поліпептидного ланцюга білка в процесі трансляції.
Що ж до реплікації вірусних ДНК, її механізм мало чим відрізняється від редуплікації генетичного матеріалу - ДНК - самої клітини. У випадку вірусних РНК реалізуються процеси, які пригнічені або зовсім відсутні в нормальних клітинах, де вся РНК синтезується тільки на ДНК як на матриці. При інфекції РНК-вірусами ситуація може бути двоякою. В одних випадках на вірусній РНК, як на матриці, синтезується ДНК ("зворотна транскрипція"), а вже на цій ДНК транскрибуються численні копії вірусної РНК. В інших, найцікавіших для нас випадках на вірусній РНК синтезується комплементарний ланцюг РНК, який і служить матрицею для синтезу – реплікації – нових копій вірусної РНК. Таким чином при інфекції РНК-вірусами реалізується принципова здатність РНК детермінувати відтворення своєї власної структури, як це має місце у ДНК.
Мультифункціональність РНК. Підсумовування та огляд знань про функції РНК дозволяють говорити про незвичайну функціональність цього полімеру в живій природі. Можна надати наступний перелік основних відомих функцій РНК.
Генетична реплікативна функція: структурна можливість копіювання (реплікації) лінійних послідовностей нуклеотидів через комплементарні послідовності. Функція реалізується при вірусних інфекціях та аналогічна головній функції ДНК у життєдіяльності клітинних організмів – редуплікації генетичного матеріалу.В цілому РНК постає перед нами настільки дивовижним полімером, що, здавалося б, ні часу еволюції Всесвіту, ні інтелекту Творця не мало б вистачити на його винахід. Як можна було бачити, РНК здатна виконувати функції обох принципово важливих життя полімерів - ДНК і білків. Не дивно, що перед наукою і постало питання: а чи не могло виникнення та самодостатнє існування світу РНК передувати появі життя в її сучасній ДНК-білковій формі?Кодуюча функція: програмування синтезу білка лінійними послідовностями нуклеотидів. Це та сама функція, що й у ДНК. І в ДНК, і в РНК одні і ті ж триплет нуклеотидів кодують 20 амінокислот білків, і послідовність триплет в ланцюзі нуклеїнової кислоти є програма для послідовної розстановки 20 видів амінокислот в поліпептидного ланцюга білка.
Структуротворча функція: формування унікальних тривимірних структур. Компактно згорнуті молекули малих РНК принципово подібні до тривимірних структур глобулярних білків, а більш довгі молекули РНК можуть утворювати і більші біологічні частинки або їх ядра.
Функція впізнавання: високоспецифічні просторові взаємодії з іншими макромолекулами (у тому числі білками та іншими РНК) та з малими лігандами. Ця функція, мабуть, основна у білків. Вона заснована на здатності полімеру згортатися унікальним чином та формувати специфічні тривимірні структури. Функція впізнавання є основою специфічного каталізу.
Каталітична функція: специфічний каталіз хімічних реакцій рибозимами. Ця функція аналогічна ензиматичної функції білків-ферментів.
ПОХОДЖЕННЯ ЖИТТЯ
Білково-коацерватна теорія Опаріна. Мабуть, першу наукову, добре продуману теорію походження життя абіогенним шляхом було запропоновано біохіміком А.І. Опарін ще в 20-х роках минулого століття [ , ]. Теорія базувалася на уявленні, що все починалося з білків, і на можливості в певних умовах спонтанного хімічного синтезу мономерів білків – амінокислот – та білковоподібних полімерів (поліпептидів) абіогенним шляхом. Публікація теорії стимулювала численні експерименти у ряді лабораторій світу, які показали реальність такого синтезу у штучних умовах. Теорія швидко стала загальноприйнятою та надзвичайно популярною.
Основним її постулатом було те, що спонтанно виникали в первинному "бульйоні" білковоподібні сполуки об'єднувалися" в коацерватні краплі - відокремлені колоїдні системи (золі), що плавають у більш розбавленому водному розчині. Це давало головну передумову виникнення організмів - відокремлення довкілля, її компартменталізацію Так як деякі білковоподібні сполуки коацерватних крапель могли мати каталітичну активність, то з'являлася можливість проходження біохімічних реакцій синтезу всередині крапель - виникала подібність асиміляції, а значить, зростання коацервату з подальшим його розпадом на частини - розмноженням. Коацерват, що асимілює, зростає і розмножується розподілом, розглядався як прообраз живої клітини (рис. 5).
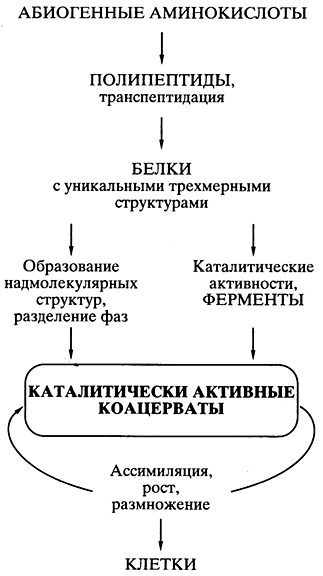
Мал. 5.Схематичне уявлення про шляхи походження життя
згідно з білково-коацерватною теорією А.І. Опаріна
Все було добре продумано та науково обґрунтовано в теорії, крім однієї проблеми, на яку довго заплющували очі майже всі фахівці у галузі походження життя. Якщо спонтанно, шляхом випадкових безматричних синтезів у коацерваті виникали поодинокі вдалі конструкції білкових молекул (наприклад, ефективні каталізатори, що забезпечують перевагу даному коацервату в зростанні та розмноженні), то як вони могли копіюватися для поширення всередині коацервату, а тим більше для передачі коацерватам- Теорія виявилася нездатною запропонувати вирішення проблеми точного відтворення - всередині коацервату і в поколіннях - одиничних ефективних білкових структур, що випадково з'явилися.
Світ РНК як попередник сучасного життя. Накопичення знань про генетичному коді, нуклеїнових кислотах і біосинтезі білків призвело до затвердження принципово нової ідеї про ТОМ, що все починалося зовсім не з білків, а з РНК [-]. Нуклеїнові кислоти є єдиним типом біологічних полімерів, макромолекулярна структура яких завдяки принципу комплементарності при синтезі нових ланцюгів (докладніше див.) забезпечує можливість копіювання власної лінійної послідовності мономерних ланок, іншими словами, можливість відтворення (реплікації) полімеру, його мікроструктури. Тому тільки нуклеїнові кислоти, але не білки можуть бути генетичним матеріалом, тобто відтворюваними молекулами, що повторюють свою специфічну мікроструктуру в поколіннях.
З низки міркувань саме РНК, а чи не ДНК, могла бути первинний генетичний матеріал.
По перше,і в хімічному синтезі, і біохімічних реакціях рибонуклеотиди передують дезоксирибонуклеотидам; Дезоксирибонуклеотиди – продукти модифікації рибонуклеотидів (див. рис. 2).
По-друге,у найдавніших, універсальних процесах життєвого метаболізму широко представлені саме рибонуклеотиди, а не дезоксирибонуклеотиди, включаючи основні енергетичні носії типу рибонуклеозид-поліфосфатів (АТФ тощо).
По-третє,реплікація РНК може відбуватися без будь-якої участі ДНК, а механізм редуплікації ДНК навіть у сучасному живому світі вимагає обов'язкової участі РНК-затравки в ініціації синтезу ланцюга ДНК.
По-четверте,володіючи всіма тими ж матричними та генетичними функціями, що і ДНК, РНК здатна також до виконання низки функцій, властивих білкам, включаючи каталіз хімічних реакцій. Таким чином, є всі підстави розглядати ДНК як пізніше еволюційне придбання - як модифікацію РНК, спеціалізовану для виконання функції відтворення та зберігання унікальних копій генів у складі клітинного геному без безпосередньої участі у біосинтезі білків.
Після того, як були відкриті каталітично активні РНК, ідея первинності РНК у походження життя отримала сильний поштовх до розвитку, і була сформульована концепція самодостатнього світу РНК,що передував сучасному життю [ , ]. Можливу схему виникнення світу РНК представлено на рис. 6.
Абіогенний синтез рибонуклеотидів та їх ковалентне об'єднання в олігомери та полімери типу РНК могли відбуватися приблизно в тих самих умовах і в тій самій хімічній обстановці, що постулювалися для утворення амінокислот та поліпептидів. Нещодавно О.Б. Четверин зі співробітниками (Інститут білка РАН) експериментально показали, що принаймні деякі полірибонуклеотиди (РНК) у звичайній водному середовищі здатні до спонтанної рекомбінації, тобто обміну відрізками ланцюга шляхом трансестерифікації. Обмін коротких відрізків ланцюга на довгі повинен призводити до подовження полірибонуклеотидів (РНК), а сама подібна рекомбінація сприятиме структурному різноманіттю цих молекул. Серед них могли б виникати і каталітично активні молекули РНК.
Навіть вкрай рідкісна поява одиничних молекул РНК, які були здатні каталізувати полімеризацію рибонуклеотидів або з'єднання (сплайсинг) олігонуклеотидів на комплементарному ланцюзі як на матриці [ ], означало становлення механізму реплікації РНК. Реплікація самих РНК-каталізаторів (рибозимів) мала спричинити виникнення самореплицирующихся популяцій РНК. Продукуючи свої копії РНК розмножувалися. Неминучі помилки в копіюванні (мутації) і рекомбінації в популяціях РНК, що самореплікуються, створювали все більшу різноманітність цього світу. Таким чином, передбачуваний древній світ РНК - це "самодостатній біологічний світ, у якому молекули РНК функціонували як генетичний матеріал, і як энзимоподобные каталізатори" .
Виникнення біосинтезу білка. Далі на основі світу РНК мало відбуватися становлення механізмів біосинтезу білка, поява різноманітних білків із успадкованою структурою та властивостями, компартменталізація систем біосинтезу білка та білкових наборів, можливо, у формі коацерватів та еволюція останніх у клітинні структури – живі клітини (див. рис. 6). ).
Проблема переходу від стародавнього світу РНК до сучасного білок-синтезуючого світу - найважча навіть для суто теоретичного рішення. Можливість абіогенного синтезу поліпептидів і білковоподібних речовин не допомагає у вирішенні проблеми, оскільки не проглядається ніякого конкретного шляху, як цей синтез міг би бути пов'язаний з РНК і підпасти під генетичний контроль. Генетично контрольований синтез поліпептидів і білків мав розвиватися незалежно від первинного абіогенного синтезу, своїм шляхом, з урахуванням вже існував світу РНК. У літературі запропоновано кілька гіпотез походження сучасного механізму біосинтезу білка у світі РНК, але, мабуть, жодна їх може розглядатися як детально продумана і бездоганна з погляду фізико-хімічних можливостей. Подаю свою версію процесу еволюції та спеціалізації РНК, що веде до виникнення апарату біосинтезу білка (мал. 7), але і вона не претендує на закінченість.
Запропонована гіпотетична схема містить два істотні моменти, які здаються важливими.
По перше,постулюється, що абіогенно синтезовані олігорибонуклеотиди активно рекомбінували за допомогою механізму спонтанної неензиматичної трансестерифікації, приводячи до утворення подовжених ланцюгів РНК і даючи початок їхньому різноманіттю. Саме цим шляхом у популяції олігонуклеотидів і полінуклеотидів могли з'явитися як каталітично активні види РНК (рибозими), так і інші види РНК зі спеціалізованими функціями (рис. 7). Більш того, неензиматична рекомбінація олігонуклеотидів, що комплементарно зв'язуються з полінуклеотидною матрицею, могла забезпечити зшивання (сплайсинг) фрагментів, комплементарних цій матриці, в єдиний ланцюг. Саме таким способом, а не каталізується полімеризацією мононуклеотидів, могло здійснюватися первинне копіювання (розмноження) РНК. Зрозуміло, якщо з'являлися рибозими, що мали полімеразну активність, то ефективність (точність, швидкість і продуктивність) копіювання на комплементарній. матриці мала значно зростати.

Мал. 7.Схема еволюції та спеціалізації молекул РНК
у процесі переходу від стародавнього світу РНК до сучасному світу
генетично детермінованого біосинтезу білків
ДругийВажливий момент у моїй версії у тому, що первинний апарат біосинтезу білка з'явився з урахуванням кількох видів спеціалізованих РНК до появи апарату ензиматичної (полімеразної) реплікації генетичного матеріалу - РНК і ДНК. Цей первинний апарат включав каталітично активну прорибосомну РНК, що мала пептидил-трансферазну активність; набір про-тРНК, що специфічно зв'язують амінокислоти або короткі пептиди; іншу прорібосомну РНК, здатну взаємодіяти одночасно з каталітичною прорібосомною РНК, про-мРНК та про-тРНК (див. рис. 7). Така система вже могла синтезувати поліпептидні ланцюги за рахунок каталізованої реакції транспептидації. Серед інших каталітично активних білків – первинних ферментів (ензимів) – з'явилися і білки, що каталізують полімеризацію нуклеотидів – реплікази, або НК-полімерази.
Втім, можливо, що гіпотеза про стародавньому світіРНК як попереднику сучасного живого світу так і не зможе отримати достатнього обґрунтування для подолання основної проблеми – науково правдоподібного опису механізму переходу від РНК та її реплікації до біосинтезу білка. Є приваблива та детально продумана альтернативна гіпотеза А.Д. Альтштейна (Інститут біології гена РАН), в якій постулюється, що реплікація генетичного матеріалу та його трансляція - синтез білка - виникали і еволюціонували одночасно і поєднано, починаючи з взаємодії абіогенно синтезованих олігонуклеотидів і аміноацил-нуклеотидилатів - змішаних ангідридів. Але це вже наступна казка... ( "І Шахразаду застиг ранок, і вона припинила дозволені промови".)
Література
1. Watson JD, Crick F.H.C. Molecular structure of nucleic acids // Nature. 1953. V. 171. P. 738-740. 2. Watson JD, Crick F.H.C. Genetic implications of structure of deoxyribose nucleic acid // Nature 1953 V. 171. P. 964-967. 3. Спірін А.С.Сучасна біологія та біологічна безпека // Вісник РАН. 1997. № 7. 4. Spirin A.S.На макромолекулярній структурі природного високополімерного рібонуклеїчного хімічного розчину // Journal of Molecular Biology. 1960. V. 2. P. 436-446. 5. Kirn S.H., Suddath F.L., Quigley GJ. та ін.Три-dimensional tertiary structure of yeast phenylalanine transfer RNA // Science. 1974. V. 185. P. 435-40. 6. Robertas J.D., Ladner J.E., Finch J.T. та ін. Structure of yeast phenylalanine tRNA at 3 A resolution // Nature. 1974. V. 250. P. 546-551. 7. Vasiliev V.D., Serdyuk I.N., Gudkov A.T., SPIRin A.S. Self-organization of ribosomal RNA // Sturcture, Function and Genetics of Ribosomes / Eds. Hardesty B. and Kramer G. New York: Springer-Verlag, 1986. P. 129-142. 8. Baserga SJ., Steitz J.A.Різноманітний світ малих ribo-nucleoproteins // The RNA World / Eds. Gesteland R.F. і Atkins J.F. New York: Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1993. P. 359-381. 9. Kruger К., Grabowski PJ., Zaug AJ. та ін. Self-splicing RNA: Autoexcision and autocyclization of the ribosomal RNA intervening sequence of Tetrahymena// Cell. 1982. V. 31. P. 147-157. 10. Guerrier-Takada С., Gardiner К., Marsh Т. та ін. RNA moiety ribonucleases P is catalytic subunit of enzyme // Cell. 1983. V. 35. P. 849-857. 11. Опарін А.І.Походження життя. М: Московський робітник, 1924. 12. Опарін А.І.Виникнення життя Землі (3-тє вид.). М: Вид-во АН СРСР, 1957. 13. Woese С. Evolution of the genetic code // The Genetic Code. New York: Harper & Row, 1967. P. 179-195. 14. Crick F.H.C. origin of the genetic code // Journal of Molecular Biology. 1968. V. 38. P. 367-379. 15. Orgel L.E. Evolution of the genetic apparatus // Journal of Molecular Biology. 1968. V. 38. P. 381-393. 16. Gilbert W. The RNA world // Nature. 1986. V 319 P. 618. 17. Joyce G.F., Orgel L.E. Prospects for understanding origin of RNA world // The RNA World / Eds. Gesteland R.F. і Atkins J.F. New York: Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1993 P 1-25. 18. Chetverina H.V., Demidenko A.A., Ugarov V.I., Chetverin A.B.Спонтанні реорганізації в RNA sequences // FEBS Letters. 1999. V. 450. P. 89-94. 19. Bartel D.P., Szostak J.W. Isolation of new ribozymes from a large pool of random sequences // Science. 1993. V. 261. P. 1411-1418. 20. Ekland E.H., Bartel D.P. RNA-catalysed RNA polymerization використовуючи nucleoside triphosphates // Nature. 1996 V. 382. P. 373-376. 21. Orgel L.E. origin of life - Review of facts and speculations // Trends in Biochemical Sciences. 1998. V. 23. p. 491-495. 22. Альтштейн А.Д.Походження генетичної системи: гіпотеза прогенів// Молекулярна біологія. 1987. Т. 21. С. 309-322.








