În toate celulele vii, proteinele sunt sintetizate de ribozomi. . Ribozomul este o macromoleculă mare cu o structură sfert asimetrică pliabilă, inspirată de acizi ribonucleici (ARN ribozomal) și proteine. Pentru a sintetiza proteine, ribozomul trebuie asigurat:
1. Un program care stabilește ordinea de ridicare a reziduurilor de aminoacizi din lancea polipeptidică a proteinei.
2. Material aminoacid, din care se va folosi proteina.
3. Energie.
Ribozomul însuși are o funcție catalitică (enzimatică), responsabilă de încorporarea legăturilor peptidice și, aparent, de polimerizarea resturilor de aminoacizi în lancea polipeptidică a proteinei.
Programul, care stabilește ordinea resturilor de aminoacizi în lancea polipeptidică a proteinei, apare sub formă de acid dezoxiribonucleic (ADN), adică din genomul clitinei. Sinteza lancelor de ARN este complementară uneia dintre lăncile de ADN i, în așa fel, confirmă exact secvența dezoxiribonucleotidică a celeilalte lance de ADN din secvența sa de ribonucleotide. Procesul de copiere a unei astfel de gene, care este modificată de enzima ARN polimerază, eliminând numele transcripției. ARN-ul în timpul sintezei acestora din urmă, în special în clitinele eucariote, poate fi supus unui număr de modificări suplimentare, numite procesare, în cursul cărora, ele pot fi modificate în părți mici ale secvenței de nucleotide. ARN, care intră, intră departe în ribozomi ca un program, care determină secvența de aminoacizi din proteina sintetizată. Se numește ARN informațional sau „mesager” (ARNm). În acest fel, însăși transcripția genelor și adoptarea ARNm asigură fluxul de informații de la ADN la ribozomi.
Materialul cob, din care va fi proteina, este aminoacizi. Cu toate acestea, aminoacizii nu sunt vicari de ribozom.Pentru a servi ca substrat pentru ribozom, aminoacidul trebuie activat pentru participarea la scindarea rezultată a ATP și acceptat (atașat covalent) de o moleculă specială de ARN, numită transfer sau transfer ARN (ARNt), pentru a ajuta la sinteza enzimelor. Aminoacil-ARNt de Otrimani este prezent în ribozom ca substrat pentru sinteza proteinelor. În plus, energia legăturii chimice dintre excesul de aminoacizi și ARNt este vicorată pentru reacția legăturii peptidice din ribozom. Astfel, activarea aminoacizilor și adoptarea aminoacil-ARNt asigură atât material, cât și energie pentru sinteza proteinelor ribozomale.
Cele trei fluxuri (informație, material și energie) sunt grupate în ribozomi. Acceptând-o, ribozomul transferă, sau traduce, informația genetică din secvența de nucleotide a ARNm la secvența secvenței de aminoacizi a proteinei lancete polipeptidice sintetizate. După cum apare în termeni moleculari, ribozomul scanează secvenţial pentru lance de ARNm (se prăbuşeşte) şi, de asemenea, selectează secvenţial din miezul aminoacil-ARNt, drept urmare specificitatea excesului de aminoacil este selectată de către ribozomul aminoacil-ARNt este determinată de specificul combinației. ARNm divizat. De asemenea, problema codului genetic este de vină: ce combinații de nucleotide determină, adică care codifică pielea cu 20 de aminoacizi, din ce molecule proteice se vor forma?
Ribozomul ARNm lancetei (sau, cu alte cuvinte, trecerea lancetei ARNm prin ribozom) stabilește o ordine timhazială strictă de intrare în ribozom a diferitelor aminoacil-ARNt, până la ordinea expansiunii combinații de nucleotide care codifică, reduc ARNm. Excesul de aminoacil al aminoacil-ARNt inversat este alăturat covalent de ribozom lancea polipeptidică în creștere. ARNt-ul deacilat este eliberat din ribozomi la situsuri. Deci secvenţial, pas cu pas, va exista o lance polipeptidică a proteinei (schema div. 1).
Biosinteza proteinelor.
Schimb plastic (asimilare și anabolism) - succesiunea reacțiilor de sinteză biologică. Numele acestui tip de schimb este ziua zilei: din discursuri, ca postura unui clitin, discursurile sunt stabilite, asemănător cu discursurile unui clitin.
Să ne uităm la una dintre cele mai importante forme de schimb plastic - biosinteza proteinelor. Biosinteza proteinelor găsit în toate proeucariotele de clitină. Informațiile despre structura primară (ordinea aminoacizilor) a unei molecule de proteine sunt codificate de secvența de nucleotide din diviziunea moleculei de ADN - genă.
Gena unei molecule de ADN, care determină ordinea aminoacizilor dintr-o moleculă de proteină. De asemenea, sub formă de nucleotide din genă, trebuie determinată ordinea aminoacizilor din polipeptidă, adică. Aceasta este structura primară, în vederea căreia toate celelalte structuri, puterea și funcția moleculei de proteine, se află în propria sa linie.
Sistemul de înregistrare a informațiilor genetice în ADN (i ARN) în secvența aparentă de nucleotide se numește cod genetic. Tobto. unitate de cod genetic (codon) - un triplet de nucleotide din ADN sau ARN care codifică un aminoacid.
Codul nostru genetic include 64 de codoni, dintre care 61 sunt codificatori și 3 sunt necodanți (codoni terminatori, care indică finalizarea procesului de traducere).
Codon-terminatori i - ARN: UAA, UAG, UGA, în ADN: ATT, ATC, ACT.
Procesul de translație este inițiat de codonul-inițiator (AUG, ADN - TAC), care codifică aminoacidul metionină. Codonul cei este primul care intră în ribozom. Apropo, se folosește metionina, deși nu se transferă, ca prim aminoacid al acestei proteine.
Codul genetic poate fi caracteristic puterii.
1. Universalitatea - codul aceluiași pentru toate organismele. Unul și același triplet (codon) în orice organism codifică același aminoacid.
2. Specificitate - codonul pielii codifică doar un aminoacid.
3. Virogenitate – mai mulți aminoacizi pot fi codificați după numărul de codoni. Există 2 aminoacizi - metionină și triptofan, care pot fi utilizați doar pentru o variantă a codonului.
4. Genele Mіzh є „semne diferite” - trei tripleți speciali (UAA, UAG, UGA), ale căror piele indică sinteza polipeptidei lanciug.
5. Nu există medii ale genei „semnelor diferite”.
Pentru a sintetiza proteina, informațiile despre secvența nucleotidelor din structura sa primară pot fi furnizate ribozomilor. Acest proces include două etape - transcriere și traducere.
Transcriere informația (rescrierea) este preluată prin sinteză pe una dintre lăncile moleculei de ADN a moleculei de ARN cu o singură lancetă, secvența nucleotidelor este exact aceeași cu secvența nucleotidelor din matrice - lancea polinucleotidică a ADN-ului .
Vaughn (i - ARN) este un intermediar care transmite informații de la ADN la locul de selecție a moleculelor proteice din ribozomi. Sinteza i ARN (transcripția) este considerată a fi un rang ofensator. Enzima (ARN - polimeraza) desparte lanceta sublinguală a ADN-ului, iar una dintre lancete (codificare) urmează principiul complementarității, nucleotidele ARN sunt vibrate. Molecula de ARN este sintetizată în acest fel (sinteza matricei) și intră în citoplasmă, iar subunități mici de ribozomi sunt înșirate la un capăt.
Un alt pas în biosinteza proteinelor este difuzat- translația tse a secvenței de nucleotide din moleculă și - ARN-ului în secvența de aminoacizi din polipeptidă. La procariote, care nu pot forma un nucleu bine format, ribozomii se pot lega la o moleculă i-ARN nou creată imediat după її divizarea ADN-ului sau se pot acumula până la finalizarea completă a sintezei її. La eucariote, i-ARN poate fi transportat în pânza de păianjen prin membrana nucleară în citoplasmă. Transferul este cauzat de proteine speciale, care formează un complex cu o moleculă de ARN. Funcțiile cremă de transfer al proteinelor citoplasmatice sunt protejate de i - ARN din bricolaj, ceea ce permite enzimelor citoplasmatice.
În citoplasmă, unul dintre capetele i - ARN-ului (și pe cel care inițiază sinteza unei molecule în nucleu) intră în ribozom și inițiază sinteza polipeptidei. În lume, trecând prin molecula de ARN, ribozomul se traduce triplet după triplet, adăugând succesiv aminoacizi la capătul în creștere al lancetei polipeptidice. Corespondența exactă a aminoacidului cu codul triplet și ARN este asigurată de t ARN.
ARN-ul de transport (ARNt) „aduc” aminoacizii în subunitatea mare a ribozomului. Molecula t-ARN poate fi pliată. Pe unele locuri dintre nucleotidele complementare se stabilesc legături de apă, iar molecula are forma unei frunze stabile. În vârful pliurilor, există un triplet de nucleotide libere (anticodon), care conține un singur aminoacid, iar baza conține un lanț de aminoacizi (Fig. 1).
Mal. 1. Schema de transport ARN: 1 - legături de apă; 2 - anticodon; 3-aminoacizi atașați.
T-ARN-ul pielii își poate transfera doar propriul aminoacid. T-ARN este activat de enzime speciale, adăugând propriul aminoacid și transportându-l la ribozom. În mijlocul ribozomilor în momentul pielii există doar doi codoni de i-ARN. Deoarece anticodonul t-ARN este complementar codonului i-ARN, există o apariție în timp util a t-ARN cu un aminoacid la i-ARN. Înainte de următorul codon vine un alt t-ARN, ca și cum ar transporta aminoacidul său. Aminoacizii sunt amestecați cu ajutorul subunității mari a ribozomului, iar cu ajutorul enzimelor se stabilesc legături peptidice împreună cu acestea. Legătura dintre primul aminoacid și її t-ARN și t-ARN merge de la ribozomul din spatele aminoacidului atacator. Ribozomul mișcă un triplet, iar procesul se repetă. În acest fel, molecula polipeptidei este construită progresiv, în care aminoacizii sunt amestecați într-o secvență de secvențe de ordinea tripleților care îi codifică (sinteza matricei) (Fig. 2).

Mal. 2. Schema pentru sinteza proteinelor: 1 – i-ARN; 2 - subunități ale ribozomului; 3 – t-ARN cu aminoacizi; 4 – t-ARN fără aminoacizi; 5 - polipeptidă; 6 - codon ta-ARN; 7-anticodon al ARNt.
Un ribozom este capabil să sintetizeze o nouă lance polipeptidică. Prote, adesea o moleculă de i-ARN prăbușește un șprot de ribozomi. Astfel de complexe se numesc poliribozomi. După terminarea sintezei, lăncile polipeptidice sunt fuzionate sub forma unei matrice - molecule i-ARN, se îndoaie într-o spirală și capătă o structură puternică (secundară, terțiară sau sfert). Ribozomii funcționează și mai eficient: prin întinderea 1c, ribozomul bacterian transformă lanceta polipeptidică din 20 de aminoacizi.
Biosinteza proteinelor (polipeptide) este un proces de pliere supra-advențios și minunat. Biosinteza proteinelor are loc în mod activ în toate organele și țesuturile, inclusiv în eritrocite. O mulțime de clitină sintetizează proteine pentru „export” (clitină hepatică, foliculi subscapulari), iar în acest fel duhoarea va răzbuna chiar și un număr mare de ribozomi. În celulele animale, numărul de ribozomi ajunge la 10 5, diametrul ribozomului ajunge la 20 nm.
Procesul de sinteză a proteinelor are loc în mijlocul celulelor de pe suprafața ribozomilor, ca complexe a două subunități cu o constantă de sedimentare de 60S și 40S, care funcționează ca o singură unitate. În ribozomi, proteinele devin 30-35%, iar ARN-ul ribozomal - 65-70%. Ribozomii au crăpături de aminoacil și peptidil. Primul servește pentru fixarea complexului de aminoacizi activi și ARNt, care este situat pe ribozom, iar celălalt fixează lancea polipeptidică, legându-se de un alt ARNt. Subunitățile ribozomilor sunt sintetizate în nucleul nucleului pe un șablon de ADN.
Esența procesului de sinteză a proteinelor este reprezentată de schema:
Sistemul de sinteză a proteinelor include ribozomi, acizi nucleici, un set de 20 de aminoacizi, diverse enzime, ATP, GTP, ioni de magneziu și aproximativ 200 de factori proteici necatalitici diferiți.
Molecula proteică este o linie lungă de reziduuri de aminoacizi, care este prezentă în formă medie de 100 până la 500 de aminoacizi. Programul pentru sinteza proteinelor pielii este stocat în molecula de acid dezoxiribonucleic (ADN). Molecula de ADN este un polimer, ai cărui monomeri sunt nucleotide. Secvența bazelor azotate dintr-o moleculă de ADN determină secvența de aminoacizi dintr-o moleculă de proteină.
Molecula de ADN are mai multe baze azotate: adenina (A), guanina (G), citozina (C) si timina (T). O secvență de trei substații (triplet) pentru a deveni un codon, care necesită un singur aminoacid.
Acizii nucleici - ADN și ARN - componente obligatorii ale biosintezei proteinelor. ADN-ul este responsabil de conservarea informațiilor genetice, în timp ce ARN-ul este responsabil de transferul acestor informații și de implementarea acesteia în apariția moleculelor de proteine. Se poate argumenta că funcția principală a ADN-ului este de a păstra genotipul, iar ARN - la genotipul virusului.
În planul clitenului, predomină ARN-ul ribozomal (ARNr). ARNr poate spiraliza trombocitele, împiedică modificarea nucleotidelor (de exemplu, 2-metil-riboză). ARNr devine aproape 80% din ARN total în clitină. Un alt tip de ARN în reprezentările clitinei de ARN de transport (ARNt), iac, iac și alte tipuri de ARN, este sintetizat în nucleu. 10-15% din cantitatea totală de ARN din clitină cade pe partea її. Au fost dezvăluite peste 60 de ARNt diferite. De asemenea, transportul altor aminoacizi se bazează pe un număr mic de ARNt diferite. Pentru aminoacizii pielii, clitinele acceptă un ARNt specific. Moleculele de ARNt sunt împărțite aleatoriu. Structurile lor au 75-93 ribonucleide.
Aminoacidul se alătură grupării 3-OH libere a mononucleotidei terminale a ARNt, reprezentată de acidul adenil. ARNt este cea mai importantă parte - un anticodon, pentru ajutorul căruia complexul de aminoacizi și ARNt este determinat de secvența a trei nucleotide din ARN-ul matricei (codon). Antidonul și codonul caută în mod complementar ajutor pentru conexiunile de apă.
Deși transportă informații despre degradare în clitin și ADN, deoarece este centralizat în nucleu, iar dacă sinteza proteinelor se realizează în citoplasmă, atunci, de asemenea, poate exista un mediator care transmite informații către citoplasma clitinei. Mediatorul cym a fost ARN mesager informațional (ARNm). ARNm este precedat de 2% din cantitatea totală de ARN de clitină. Molecule de ARNm găsite (cuprind până la 5 mii de nucleotide). ARNm conține, de asemenea, baze azotate. Trei dintre ele (A, G, C) sunt la fel ca în ADN, iar al patrulea este uracil.
Informația codificată în ARNm este necesară pentru sinteza unei molecule de proteine care se găsește pe ribozomi. Sinteza ARNm în nucleul celulelor este destul de lentă, ceea ce este necesar pentru biosinteza activă a moleculelor de proteine. ARNm se stabilește pe una dintre catenele de ADN din nucleu. În acest caz, structura dublu catenară a ADN-ului se desfășoară și, cu participarea ARN polimerazei epuizate de ADN, sinteza ARNm urmează principiul complementarității:

Schema pentru sinteza ARNm
Principiul complementarității înseamnă că adenina de pe helixul ADN-ului este asociată cu ARNm de uracil, timina cu adenină și guanina cu citozină. De asemenea, ARNm citește informații din ADN.
Stadiul ADN - ARN, în acest fel, semnifică sinteza moleculei de ARNm, în care secvența de nucleotide este complementară cu diviziunea cântecului (gena) ADN-ului. Acest proces se numește transcripție. Apoi ARNm este situat pe ribozom, unul câte unul cu subunități її. O moleculă de ARNm este fixată pe mulți ribozomi în același timp, formând așa-numiții polizomi. Prezența polizomilor promovează eficiența și flexibilitatea vicariozității ARNm.
Sinteza lancetei polipeptidice a depozitului de variante se realizează pe matricea ARNm. Procesul de transfer al ARNm la o proteină prin omiterea numelui traducerii. Etapa „ARN -> proteină” este procesul de sinteză a proteinelor, care dirijează ARNm. În acest mod, transferul de informații zavzhdi merge direct ADN - ARN - proteină.
Procesul de traducere include următorii pași:
Aceeași enzimă participă la fixarea unui aminoacid activat înainte în poziția 2 sau 3 a ribozei nucleotidei ARNt rămase:
În acest complex de aminoacizi, ribozomul este transportat și are loc sinteza unei molecule de proteine. Aminoacil-ARNt sintetaza este specifică, poate recunoaște ca aminoacid, i-ARNt. În clitină, în această ordine, există cel puțin 20 de sintetaze diferite, variind până la numărul de a-aminoacizi.
2. ARNt, legat printr-o legătură eterică cu un singur aminoacid, se localizează pe ribozom și se interacționează cu ARNm pentru tipul de complementaritate între un triplet specific de nucleotide ARNm, numele unui codon și tripletul nucleotid specific complementar (anticodon) tr. În acest fel, codonul de piele ARNm asigură fixarea specifică a unui aminoacid în lancea peptidică pentru ajutorul anticodonului ARNt. Ribozomul trece peste moleculele de ARNm, citind secvenţial toţi codonii, stabilindu-se astfel ordinea dizolvării tuturor aminoacizilor care sunt livraţi la locul de sinteză.
Sinteza moleculei proteice trece direct de la gruparea amino liberă la gruparea carboxil liberă a aminoacidului. Denumirea aminoacidului din sinteza lancetei polipeptidice este metionina, pentru care codonul este secvența de nucleotide a ARNm AUG.
Inițierea sintezei polipeptidelor începe cu fixarea a doi anticodoni ARNt în spatele celui de-al doilea codon ARNm. Procesul de creștere a vizibilității energiei, care trebuie să servească drept GTP, precum și participarea unui număr de factori proteici inițiatori și peptidiltransferaza.
Pentru participarea acestei enzime, viteza iluminării legaturi covalente ajunge la 1200 de aminoacizi / xv / ribozomi.

Schema de inițiere a sintezei polipeptidelor
3. După ce dipeptida „nedorită” este introdusă, ARNt-ul umple ribozomul și poate furniza noi molecule de aminoacizi, iar ARNm este împins prin ribozomi (polizomi) de către trei nucleotide. Ca rezultat al relocarii (translocării), codonul liber ocupă o poziție de recunoaștere a moleculei de ARNt de bază. De asemenea, în stadiul de alungire, există o secvență de adăugare a unui aminoacid la lanceta polipeptidică în ordinea codonilor moleculei de ARNm.
Lancea polipeptidică, care se presupune că este atașată la o moleculă de ARNt, este fixată de marea subunitate a ribozomului. Adăugarea de aminoacid suplimentar dermal la lancea polipeptidică se datorează relației dintre gruparea amino a aminoacidului, care se unește în complexul cu ARNt și gruparea carboxil a peptidei.
4. Terminarea sau finalizarea sintezei moleculei polipeptidice prin obținerea terminației codonului „fără sens” și a factorului de terminare a proteinei. Există trei codoni (UAH, UGA, UAA), care nu codifică, nu se leagă de un aminoacid, deoarece nu există anticodoni în ARNt complementari acestora în celule. Teoretic, un singur codon „fără sens”, care este responsabilitatea sintezei moleculei proteice, este responsabil pentru trecerea polizomului a 5-3 ARNm.
Prezența unui codon de terminare în orice diviziune a ARNm înseamnă terminarea sinteza proteinei. Ca rezultat, polizomul se dezintegrează, ARNm nevicoristan este hidrolizat de polinucleozid fosforilază, iar subunitățile ribozomului sunt pregătite pentru a începe sinteza unei noi molecule de proteine.
ARNm poate participa în mod repetat la procesul de biosinteză a proteinelor. Trivalitatea funcționării moleculei de ARNm nu este aceeași în diferite organisme. Vaughn se poate legăna de la câteva fluiere la câteva dib.
5. În ADN, structura primară a proteinei este codificată. Prin urmare, sintetizarea pe ribozomii moleculei de proteină poate fi încă o stare completă reziduală. Ele reprezintă polipeptide primare, care recunosc apoi modificări numerice (asocierea monomerilor cu oligomeri stabili, adăugarea de coenzime, transformări chimice) care modifică structura proteinelor și, prin urmare, activitatea acestora.
Structurile secundare și terțiare nu sunt codificate, ele sunt determinate de puterea structurii primare și înseamnă că această formă chi insha a moleculei proteice este depusă în secvența de aminoacizi și posibilitățile interacțiunilor lor între ele. Modificările structurale ale proteinelor, care sunt sintetizate, pot fi mai frecvente la ribozomi egali sau după terminarea sintezei ca urmare a adăugării altor grup functional.
M-am uitat la schema de transmisie la vedere
te poți schimba în moduri diferite. Deci, în viruși, care nu răzbună ADN-ul, informația este încorporată în ARN. Când virusul pătrunde în clitină, informația este transferată în ADN-ul clitinei, iar restul sintetizează deja ARNm, pe care proteinele virusului sunt sintetizate pe matrice. Un astfel de proces se numește transcriere inversă, iar schema de transmitere în orice fel va fi ofensatoare:
Atâta timp cât secvența de nucleotide din ADN și, prin urmare, ARNm este păstrată, natura proteinei, care este nou sintetizată, rămâne neschimbată.
Informația genetică necesară pentru sinteza proteinelor poate fi prezentată în mod similar cu înregistrarea limbajului uman, deoarece este formată din succesiunea de litere care formează cuvintele acelui discurs. În limbajul genetic, însă, există mai puține litere chotiri - baze chotiri (adenină, guanină, uracil, citozină).
Codul genetic include cuvinte triliterale. Chotiri sugerează în acest fel (43) să dea 64 de opțiuni (cuvinte), care sunt mai mult sau mai puțin suficiente, pentru a codifica 20 de aminoacizi. În această ordine, 64 de codoni alcătuiesc codul genetic (Tabelul 3).
Analiza codului genetic arată că numărul de codoni este diferit pentru diferiți aminoacizi. De exemplu, metionina și triptofanul pot avea un singur codon, în timp ce arginina, leucina și serina pot avea șase codoni fiecare. Prezența unui număr de codoni pentru un aminoacid reflectă „virogeneitatea” codului. De asemenea, acel aminoacid în sine poate fi codificat ca un kilkom pentru propriile sale triplete de nucleotide. În același timp, tripletul pielii conține un aminoacid întreg în lanceta polipeptidică, care este sintetizat.
Tabelul 3
cod genetic
|
nucleotide |
O altă nucleotidă |
nucleotide |
|||
Codul genetic este universal și același la speciile de dezvoltare diferită (oameni, creaturi, plante, microorganisme). Universalitatea codului este de a spune că toate organismele vii din trecutul mic strămoș.
Aminoacizii okremi (hidroxiprolină, oxilizină), de exemplu, nu se încurcă cu codonul și se mulțumesc cu ajutor reacții chimice deja după sinteza polipeptidei lanceug. Acest proces, luând numele de modificare post-traducțională și chiar mai important pentru buna funcționare a proteinei pielii.
Codonii tăcuți (UAA, UAG, UGA) nu codifică aminoacizi, proteina este într-adevăr un semnal pentru finalizarea sintezei unei molecule de proteine.
Astfel, ARNm este un purtător neîntrerupt de informații genetice de la nucleu la ribozomul citoplasmei. Un ribozom împrumută aproximativ 80 de nucleotide per ARNm și catalizează aproximativ 100 de legături peptidice per quilină (Severin E. S. și colab., 2011).
Sinteza moleculelor de proteine poate fi supusă modificărilor structurale pe ribozomi egali sau după finalizarea sintezei ca urmare a adăugării diferitelor grupări funcționale. ARNm citoplasmatic poate avea o perioadă relativ scurtă. Cantitatea de ARNm este sintetizata si stocata in forma inactiva, fiind pregatita pentru sinteza rapida a proteinelor. Informațiile ARNm Oscilki sunt legate de secvența liniară a nucleotidelor, integritatea secvenței este foarte importantă. Dacă o modificare a ordinii nucleotidelor poate modifica sau nu sinteza proteinei. Anul acesta a fost stabilit un număr redus de inhibitori ai replicării ADN-ului în organismele clitinice (antibiotice, vidre chimice, medicamente antivirale). Modificările în secvența bazelor purinice sau pirimidinice din genă au luat numele mutației.
Înlocuirea a mai mult de o nucleotidă într-un codon (mutație) duce la o schimbare în codificarea unui aminoacid pentru altul. De exemplu, o mutație datorată înlocuirii acidului glutamic cu valină în molecula de hemoglobină duce la sinteza hemoglobinei, care duce la anemie asemănătoare secerului. Astăzi, au fost raportate peste 200 de mutații ale polipeptidei cola a moleculei de hemoglobină umană. Adesea, mutagenii sunt vorbirea (nitrozaminele, de exemplu), care modifică structura bazelor azotate, ceea ce duce la o schimbare a naturii complementarității bazelor. Reflexia ultraviolete a provocat condensarea excesului de timină cu dimerii de timină dizolvați. Din fericire, în urma afluxului dur de schimbări ultraviolete, creaturile sunt protejate de balonul de ozon al atmosferei.
O mulțime de antibiotice, care se găsesc în practica veterinară, inhibă sinteza proteinelor bacteriene (lincomicina, eritromicină, cloramfenicol) în stadiul de traducere. Cu această clitina microbiană, gyne chi își crește dezvoltarea. Astfel de antibiotice, cum ar fi tetraciclinele, nu interferează cu sinteza ribozomală la viețuitoare. Penicilinele nu sunt inhibitori direcți ai sintezei proteinelor, ele acționează ca inhibitori ai bacteriilor prin blocarea sintezei hexapeptidelor în peretele celular. Trebuie remarcat faptul că sinteza proteinelor are loc nu numai pe ribozomi, ci și în mitocondrii. Mitocondriile pot avea un aparat independent de sinteză a proteinelor pentru nevoile lor, deși nu toate proteinele mitocondriale sunt sintetizate în aceste organite. ARN-ul mitocondrial devine mai puțin de 3% din cantitatea de ARN de clitină. Ribozomii mitocondriilor sunt mai mici în spatele marginilor, mai mici sunt citoplasmatici. Codonul UGA, ca terminator pentru sinteza proteinelor în citoplasmă, se găsește în mitocondrii lângă codonul UGG pentru codificarea aminoacizilor.
Sinteza proteinelor pe ribozomi nu creează un corp complet rezidual. Ele reprezintă polipeptidele primare, care recunosc apoi modificări numerice (asocierea monomerilor cu oligomeri, adăugarea de coenzime, transformări chimice) care modifică structura proteinei și, prin urmare, activitatea acesteia.
Imaginea 9 din prezentarea „Biosinteza proteinelor” la lecții de biologie pe tema „Biosinteza proteinelor”
Dimensiune: 960 x 720 pixeli, format: jpg. Pentru a descărca gratuit poza pentru lecția de biologie, faceți clic pe imagine cu butonul din dreapta al mouse-ului și apăsați „Salvați imaginea ca...”. Pentru a afișa imagini la lecție puteți descărca și prezentarea „Biosinteza proteinei.pptx” cu toate pozele în zip-arhivă. Arhiva Rozmir - 1719 KB.
Fa o prezentare„Funcțiile albilor” – Așa sunt percepute recepția semnalelor din mediul exterior și transmiterea informațiilor către client. Odată cu descompunerea a 1 g de proteină în produsele finale, se observă 17,6 kJ. Ce este renaturarea? Podіb'єmo pіdbags: 9. Catalitic. Procesul de refacere a structurii proteinei după denaturare se numește renaturare. Pimenov A.V. Proteinele sunt una dintre sursele de energie din celule.
„Proteinele vorbirii” – De exemplu: colagen. Biologie Vikladach: Boldireva L.A. . fund: ou fiert. Amino acid - vorbire organică, Proteine nerezolvate - fibrilare. albii Harchov. . proteine zahisnі. Structura proteinei. Vikoristovuyutsya organism pentru ruhu. Proteine energetice.
„Proteine și funcții yoga” – Rol catalitic. Funcția motorului. Conceptul de proteine. Hidroliza proteinelor duce la scindarea legăturilor polipeptidice: Visnovok: Vasele purtătoare de sânge, tendoanele și părul au fost induse din proteine. Budova și funcțiile proteinei. Putere chimică bilkiv. proteinele participă la membranele stabilite de clitină, organoizi și membranele de clitină.
„Biosinteza proteinelor” – Lista literaturii. Introducere. 4. Zmist. Biosinteza proteinelor în celulele vii. 7. 10. 9. Schema clitinei în creștere și viață. 5. 6. 1. 8. 2. 3.
„Biosinteza proteinelor” - Traducere (lat. Transfer, traducere). Transcriere (lat. rescriere). Întoarce-te. Înţeles whites. Zmist. Energetica biosintezei. Rolul enzimelor. Sinteza polipeptidei lanciug pe ribozomi. 5. Care este secvența nucleotidelor din i-ARN înregistrată pe crucea ADN: T-A-C-G-G-A-T-C-A-C-G-A -G-T-G-C-T A-U -G-Ts-G-U-A-G-U-G-Ts-U A-U-G-Ts-Ts-UG
„Biosinteza biologiei proteinelor” - Mikola Kostyantinovich Koltsov (1872-1940). AG Funcția principală a ribozomilor este sinteza proteinelor. Dogma centrală (postulatul de bază) al biologiei moleculare este sinteza matricei. C. Anticodon - un triplet de nucleotide pe partea superioară a ARNt. Biosinteza proteinelor. După terminarea sintezei, iARN se descompune în nucleotide.
Temele au 8 prezentări în total
© A.S. Spirin
BIOSINTEZ BILKIV, SVIT ARNLA FEL DE. Spirin
Spirin Oleksandr Serghiovici– academician, director al Institutului de Biologie al Academiei Ruse de Științe, membru al Prezidiului Academiei Ruse de Științe.
Mai 1953, D. Watson și F. Crick au descoperit principiul organizării structurale (moleculare) a vorbirii genelor - acidul dezoxiribonucleic (ADN). Structura ADN-ului a dat cheia mecanismului de creare exactă - reduplicare - a vorbirii genelor. Deci vinicla este o nouă știință - biologia moleculară. Bulo a formulat așa-numita dogmă centrală a biologiei moleculare: ADN Yu ARN Yu proteine. Sensul se bazează pe faptul că informația genetică este scrisă în ADN, este realizată în proteine, dar nu fără un punct de mijloc, ci în spatele unui polimer suplimentar de spori - acidul ribonucleic (ARN), iar calea de la acizi nucleici la proteine nu este limitată. . În acest fel, ADN-ul este sintetizat pe ADN, asigurând puterea de reduplicare, astfel încât să se creeze materialul genetic rămas în generații; ARN-ul este sintetizat pe ADN, rezultând rescriere, transcriere, informații genetice sub formă de copii numerice ale ARN; Moleculele de ARN sunt modele pentru sinteza proteinelor - informația genetică este tradusă sub formă de lănci polipeptidice. În cazuri speciale, ARN-ul poate fi rescris sub formă de ADN („transcripție inversă”) și, de asemenea, copiat sub formă de ARN (replicare), dar proteinele nu pot fi niciodată un șablon pentru acizi nucleici (raport div.).
De acum înainte, însuși ADN-ul semnifică declinul organismelor, astfel încât recrutarea proteinelor, care se găsesc în generații, și semnul asociat acestora. Biosinteza proteinelor este procesul central al materiei vii, iar acizii nucleici sunt responsabili de acest lucru, pe de o parte, cu un program care determină întregul set și specificul proteinelor care sunt sintetizate și, pe de altă parte, prin mecanism. a unor programe precise de replicare în generaţii. De acum înainte, speranța de viață în forma clitinică modernă duce la justificarea mecanismului de biosinteză redusă a proteinelor.
BIOSINTEZ BILKIV
Dogma centrală a biologiei moleculare postulează doar modalități de a transfera informații genetice de la acizi nucleici la proteine și, apoi, la putere și acel semn al unui organism viu. Dezvoltarea mecanismelor pentru implementarea acestei căi timp de un deceniu, care a urmat formularea dogmei centrale, a extins bogat diferite funcții ale ARN-ului, dar este doar un purtător de informații de la gene (ADN) la proteine și servire și o matrice. pentru sinteza proteinelor.
Pe fig. 1 prezintă o schemă generală a biosintezei proteinelor în clitină. ARN mesager(ARN mesager, ARN mesager, ARNm), care codifică proteine, așa cum s-a spus mai sus, este doar una dintre cele trei clase principale de ARN-uri celulare. Masa principală їhnyu (aproximativ 80%) pentru a deveni următoarea clasă de ARN - ARN ribozomal, yakі utvoryuyut cadrul structural și centrele funcționale ale particulelor universale de sinteză a proteinelor - ribozomi. ARN-ul ribozomal în sine este viabil - atât structural, cât și funcțional - modelând mașini moleculare ultramicroscopice numite ribozomi. Ribozomii preiau informații genetice precum moleculele de ARNm și, fiind programați de restul, încolțesc proteine în modelul exact conform programului.
Prote, schob sintetizează proteine, lipsite de informații și programele sunt insuficiente - consumatoare de material, din care se poate lucra. Material potic pentru sinteza proteinelor în ide în ribozomi pentru ajutorul clasei a treia de ARN celular. transfer ARN(ARN de transfer, ARN de transfer, ARNt). Duhoarea leagă covalent - acceptă - aminoacizii, ca material în devenire pentru proteine și pare că aminoacil-ARNt se găsește în ribozom. În ribozomi, aminoacil-ARNt interacționează cu codoni - combinații de trinucleotide - ARNm, după care decodificarea codonilor are loc în timpul procesului de traducere.
Acizi ribonucleici
De asemenea, este posibil să se recruteze principalul ARN celular, principalul proces principal al materiei vii moderne - biosinteza proteinelor. ARNm Ce, ARN ribozomal și ARNt. ARN-ul este sintetizat pe ADN cu ajutorul enzimelor - ARN polimeraze, care induce transcripția - rescrierea diviziunilor liniare (brațele liniare) ale ADN-ului dublu catenar în formă de ARN monocatenar. Celulele ADN-ului, care codifică proteine celulare, sunt rescrise sub formă de ARNm, precum și pentru sinteza de copii numerice ale ARN-ului ribozomal și ARNt - celule speciale ale genomului clitinei, pentru care există o rescriere intensivă fără traducere ulterioară a proteina.
Structura chimică a ARN-ului.
Din punct de vedere chimic, ARN-ul este deja similar cu ADN-ul. Discurs ofensator - polimeri liniari întregi de nucleotide. Monomerul pielii - o nucleotidă - este o fosforilare a N-glicozidei, determinând un exces de inel cu cinci atomi de carbon - pentoză, care poartă o grupare fosfat pe gruparea hidroxil a celui de-al cincilea atom de carbon (legătură de pliere) și o bază azotată la primul atom de carbon (N-glicozină) Diferența chimică principală dintre ADN și ARN în ceea ce are un exces de monomer ARN - ceriboză și monomerul ADN - deoxiriboză, care este similar cu riboza, are o grupare hidroxil zilnică cu o altă atom de carbon (fig. 2).
 Mal. 2. Formule chimice ale stocurilor una dintre ribonucleotide - uridil acid (U) și youmu omolog dezoxiribonucleotide - acid timidilic (dT) |
Baze azotate i în ADN, i în ARN chotiri vide: două baze purinice - adenină (A) și guanină (G) - două baze pirimidinice - citozină (C) și uracil (U) sau timină yogo metilata (T). Uracilul este caracteristic monomerilor ARN, iar timina este caracteristică monomerilor ADN, iar ARN și ADN-ul sunt la fel de importante. Monomerii - ribonucleotidele ARN și dezoxiribonucleotidele ADN - formează o lance polimerică pentru formarea suplimentară de situsuri fosfodiester între surplusurile de sânge (între cei cinci și al treilea atom de carbon ai pentozei). Astfel, lancea polimerică a acidului nucleic - ADN sau ARN - poate fi reprezentată ca un schelet liniar zahăr-fosfat cu baze azotate ca grup biologic. Structura macromoleculară a ARN-ului. Diferența macrostructurală fundamentală dintre cele două tipuri de acizi nucleici se datorează faptului că ADN-ul este o singură spirală, adică o macromoleculă cu două legături ale catenelor polimerice complementare, răsucite spiralat în jurul axei centrale (div. [ , ]), și ARN - polimerul monocatenar. În același timp, interacțiunile reciproce ale grupurilor biologice - baze azotate - una câte una, precum și cu fosfații și hidroxizii din coloana vertebrală zahăr-fosfat sunt aduse în punctul în care polimerul ARN monocatenar se învârte pe sine și se răsucește în o structură compactă, similară cu plierea unei lancete polipeptidice a unei proteine într-un globule compact. În acest fel, secvențele unice de nucleotide ale ARN pot forma întinderi unice de structuri. |
Anterior, structura spațială specifică a ARN a fost demonstrată la descifrarea structurii atomice a unui ARNt în 1974. [ , ] (Fig. 3). Formarea unei lancete polimerice de ARNt, care constă din 76 de monomeri nucleotidici, duce la formarea unui miez globular compact arcuit, din care două creste sunt șterse cu o tăietură dreaptă. Mirosurile sunt spirale scurte de subsârmă pe clusterul de ADN, dar sunt, de asemenea, organizate cu ajutorul sprijinului reciproc al unuia și aceluiași tієї și ARN lancet. Unul dintre ele acționează ca acceptor de aminoacizi și participă la sinteza proteinei lancete polipeptidice pe ribozom, iar celălalt este utilizat pentru interacțiunea complementară cu tripletul ARNm (codonul) care codifică în același ribozom. Doar o astfel de structură este concepută special pentru a interacționa cu proteina-enzimă, care atârnă aminoacidul de ARNt și ribozomul în procesul de traducere, astfel încât să fie „recunoscut” în mod specific de către aceștia.

Mal. 3. Modele atomice (stângaci) și scheletice (dreaptaci) de drojdie de ARNt de fenilalanină
Producția de ARN-uri ribozomale izolate a dat o aplicație ofensivă și expansivă pentru formarea de structuri specifice compacte ale polimerilor liniari încă mai mari de același tip. Ribozomul este alcătuit din două părți nervoase - subpărți (subunități) ribozomale mari și mici. Subparticula de piele a fost inspirată de un ARN polimeric înalt și o serie întreagă de diferite proteine ribozomale. Lungimea lancelor în ARN-ul ribozomal este semnificativă: de exemplu, ARN-ul subunității mici a ribozomului bacterian este de peste 1500 de nucleotide, iar ARN-ul subunității mari este de aproximativ 3000 de nucleotide. La salvatori, inclusiv la oameni, ciARN-ul este chiar mai mare - aproape de 1900 de nucleotide și peste 5000 de nucleotide în subpărți mici și mari, aparent.
S-a demonstrat că ARN-ul ribozomal, apa-argint este izolat de partenerii săi proteici și îndepărtat din aspect curat, structurile în sine se pliază spontan într-o structură compactă, similară ca mărime și formă cu subunitățile ribozomale]. Forma subparticulelor mari și mici este diferită, iar forma ARN-urilor ribozomale mari și mici este similară (Fig. 4). În acest fel, lăncile liniare ale ARN-ului ribozomal se auto-organizează într-o întindere specifică de structură, ceea ce înseamnă expansiune, formă i, eventual, atașarea internă a subpărților ribozomiale și, de asemenea, toți ribozomii.
ARN minor. În lume, a fost stabilită producția de componente în clitină vie și celelalte trei fracții din ARN-ul total al clitinei, dar trei dintre principalele tipuri de ARN din dreapta nu sunt amestecate. S-a dovedit că în natură există alte tipuri impersonale de ARN. Acesta este motivul pentru care îl numim „ARN mic”, care poate acoperi până la 300 de nucleotide, adesea cu funcții necunoscute. De regulă, mirosurile sunt asociate cu una sau mai multe proteine dekilcom și sunt prezentate în termenul de ribonucleoproteine - „RNP mic”.
ARN-urile mici sunt prezente în toate celulele clitinei, inclusiv în citoplasmă, nucleu, nucleu, mitocondrii. Majoritatea acestor RNP-uri mici, care funcționează într-un fel, participă la mecanismele de procesare post-transcripțională a principalelor tipuri de ARN (prelucrare ARN) - transformarea precursorilor ARNm pe ARNm matur (splicing), redagularea ARNm, biogeneza ARNt. , maturarea ARN-ului ribozomal. Una dintre cele mai abundente reprezentări ale speciilor mici de RNP (SRP) în clitină joacă un rol cheie în transportul proteinelor care sunt sintetizate prin membrana clitinei. Vіdomi vidi ARN mic, funcțiile de reglementare scho vykonuyut în traducere. Un mic ARN special intră în depozitul celei mai importante enzime, care este cunoscută pentru promovarea reduplicării ADN-ului în generațiile de clitină - telomeraza. Inutil să spunem că dimensiunile lor moleculare pot fi comparate cu dimensiunile proteinelor globulare celulare. În acest mod, pas cu pas, devine clar că funcționarea celulelor vii nu este atribuită numai diferitelor proteine, care sunt sintetizate într-un set bogat de ARN-uri diferite, ci și ARN mic nіst i razmiri bіlkіv.
Ribozima. Toată viața activă este motivată de schimbul vorbire - metabolism, iar toate reacțiile biochimice ale metabolismului sunt dependente de resursele necesare pentru siguranța vieții doar de catalizatori specifici extrem de eficienți, crearea evoluției. Actualizarea bogăției unui deceniu de biochimie s-a dovedit că cataliza biologică este în mod constant și peste tot fiind numită proteine, așa cum sunt numite. enzime, sau enzime. I axa y 1982-1983 pp. S-a demonstrat că în natură є specii de ARN, care, similar proteinelor, pot avea activitate catalitică foarte specifică [ , ]. Astfel de catalizatori ARN au fost numiți ribozime. Declarația despre vinyatkovistitatea proteinelor în cataliza reacțiilor biochimice a luat sfârșit.
În zilele noastre, ribozomul este, de asemenea, privit ca o ribozimă. Într-adevăr, toate datele experimentale recente indică faptul că sinteza proteinei lanceolate polipeptidice în ribozom este catalizată de ARN-ul ribozomal, și nu de proteinele ribozomale. A fost identificată o celulă catalitică a marelui ARN ribozomal, care este responsabilă de catalizarea reacției de transpeptidare, care ajută la creșterea proteinei lancete polipeptidice în procesul de translație.
În ceea ce privește replicarea ADN-ului viral, acest mecanism este puțin explorat în reduplicarea materialului genetic - ADN - al celulei în sine. În cazul ARN-urilor virale se realizează procese care sunt suprimate sau uneori zilnic în celulele normale, unde tot ARN-ul este sintetizat doar pe ADN, ca pe o matrice. Când sunt infectați cu viruși ARN, situația poate fi dublă. În unele cazuri, pe ARN-ul viral, ca pe un șablon, ADN-ul este sintetizat („transcripție inversă”), iar copiile numerice ale ARN-ului viral sunt transcrise pe acest ADN. În alte atacuri, cele mai importante pentru noi asupra ARN-ului viral, este sintetizată o lancetă de ARN complementară, care servește ca șablon pentru sinteza - replicarea - de noi copii de ARN viral. În acest fel, în timpul infecției cu virusuri ARN, principiul formării ARN-ului este realizat pentru a determina crearea propriei structuri de apă, așa cum este cazul ADN-ului.
Multifuncționalitatea ARN-ului. Având în vedere cunoștințele despre funcțiile ARN, este posibil să vorbim despre funcționalitatea non-primară a acestui polimer în natura vie. Este posibil să se dea o schimbare ofensivă a principalelor funcții ale ARN.
Funcția replicativă genetică: capacitatea structurală de a copia (replica) secvențe liniare de nucleotide prin secvențe complementare. Funcția se realizează în infecțiile virale și este similară cu funcția principală a ADN-ului în viața organismelor clitinice - reduplicarea materialului genetic.În ansamblu, ARN-ul stă în fața noastră cu un polimer minunat, care, n-ar fi fost, nici pentru timpul evoluției Lumii-Toate, nici pentru intelectul Creatorului, nu ar fi suficient să vorbim despre vin de yoga. Deoarece este posibil să bulo bachiti, ARN-ul este capabil să încalce funcțiile atât ale vieții fundamentale ale polimerilor - ADN-ul și proteinele. Nu este de mirare că știința a primit hrană: de ce nu ar putea acea fundație autosuficientă a lumii ARN-ului să fie distrusă de apariția vieții în forma sa modernă de ADN-proteine?Funcția de codificare: programarea sintezei proteinelor prin secvențe liniare de nucleotide. Aceasta este însăși funcția pe care o are ADN-ul. I în ADN, i în ARN, unul și același triplet de nucleotide codifică 20 de aminoacizi ai proteinei, iar secvența tripletului din lăncile acidului nucleic este un program pentru ordonarea secvențială a 20 de tipuri de aminoacizi în lancea polipeptidică a proteinei.
Funcția de creare a structurii: formarea unor structuri triviale unice. Moleculele mici de ARN pliate compact sunt practic similare cu structurile trivimer ale proteinelor globulare, iar moleculele de ARN mai mari pot forma particule sau nuclei biologici mai mari.
Funcția de recunoaștere: interacțiuni spațiale foarte specifice cu alte macromolecule (inclusiv proteine și alți ARN) și liganzi mici. Această funcție, poate, este cea principală în proteine. Se bazează pe zdatnostі polimer zgortatisya rang unic și modelarea structurilor trivimіrnі specifice. Funcția de recunoaștere este baza catalizei specifice.
Funcția catalitică: cataliză specifică a reacțiilor chimice de către ribozime. Această funcție este similară cu funcția enzimatică a proteinelor-enzime.
TRĂIND VIAȚA
Teoria protein-coacervat a Oparinului. Poate, știința pershu, o teorie bine gândită a călătoriei vieții pe o cale abiogenă a fost sugerată de biochimistul A.I. Oparin în anii 20 ai secolului trecut [ , ]. Teoria s-a bazat pe faptul că totul a pornit de la proteine și pe posibilitatea sintezei chimice spontane a monomerilor proteinelor - aminoacizi - și a polimerilor asemănători proteinelor (polipeptide) în mod abiogen. Publicarea teoriei a stimulat experimente numerice într-un număr de laboratoare din lume, deoarece acestea au arătat realitatea unei astfel de sinteze în mințile piese. Teoria lui Shvidko a devenit sălbatic acceptată și extrem de populară.
Principalul postulat її au fost cei care au blamat în mod spontan în „bulionul” primar, plăcile asemănătoare proteinelor au fost combinate „în picături coacervate - coloane armate cu apă ale sistemului (soluri), care pluteau în apropierea golului de apă diluat mai mare. nennya organіzmіv - vіdokremlenny dovkilla, її compartimentare Deoarece proprietățile asemănătoare proteinei ale dungilor coacervate ar putea provoca activitate catalitică, a fost posibil ca reacțiile biochimice să treacă prin sinteza în mijlocul dungilor - simi vinice , creșterea coacervatului cu dezintegrarea yogo la distanță în părți - reproduceri . Coacervatus, care se asimilează, crește și se înmulțește sub piele, arătând ca un prototip al unei celule vii (Fig. 5).
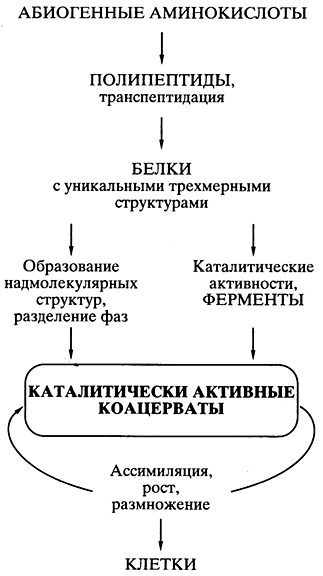
Mal. 5. Reprezentarea schematică a căilor călătoriei vieții
conform teoriei proteine-coacervate a A.I. Oparina
Totul a fost bine gândit și întemeiat științific în teorie, cu excepția unei probleme, multă vreme ochii tuturor fahivților din galerie au fost turtiți mult timp. Ca și cum spontan, de-a lungul traseului sintezelor fără matrice în coacervat, construcțiile moleculelor proteice (de exemplu, catalizatori eficienți, care asigură predominarea acestui coacervat în creșterea reproducerii) au fost vibrate una câte una în depărtare, apoi deoarece duhoarea ar putea fi copiată pentru a se extinde în coacervatele mijlocii și mai mult pentru transmiterea la coacervate - Teoria a apărut intempestiv pentru a propaga problema creării exacte - în mijlocul coacervatului și în generații - a structurilor proteice eficiente unice, care a apărut din senin.
ARN ușor ca front viata de zi cu zi. Cunoștințe acumulate despre cod genetic, acizii nucleici și biosinteza proteinelor au condus la confirmarea unei idei fundamental noi despre TOM, că totul a pornit nu de la proteine, ci de la ARN [-]. Acizii nucleici sunt singurul tip de polimeri biologici, a căror structură macromoleculară urmează principiul complementarității în sinteza noilor lănci (raport div.) asigurând posibilitatea copierii secvenței liniare umede a lankurilor monomerice, cu alte cuvinte, posibilitatea de crearea (replicarea) polimerului, microstructura acestuia. Prin urmare, numai acizii nucleici, dar nu și proteinele, pot fi un material genetic, adică molecule care își repetă microstructura specifică în generații.
Nivelul scăzut de ARN în sine, și nu ADN-ul, ar putea fi primul material genetic.
Primul, i în sinteza chimică, i în reacțiile biochimice, ribonucleotidele sunt transformate în dezoxiribonucleotide; Dezoxiribonucleotidele sunt produse ale modificării ribonucleotidelor (div. Fig. 2).
Într-un mod diferitîn cele mai recente, procese universale ale metabolismului vieții, ribonucleotidele, mai degrabă decât dezoxiribonucleotidele, sunt larg reprezentate, inclusiv principalele surse de energie de tip ribonucleozid-polifosfat (ATP lean).
În al treilea rând, Replicarea ARN poate fi efectuată fără nicio implicare a ADN-ului, iar mecanismul reduplicării ADN-ului poate fi găsit în lumea vie pentru a influența rolul general al primerului ARN în inițierea sintezei lancei ADN.
În al patrulea rând care posedă toate aceleași matrice și funcții genetice, cum ar fi ADN-ul, ARN-ul, sunt, de asemenea, capabili de funcții scăzute, proteine puternice, inclusiv catalizarea reacțiilor chimice. În acest fel, toți considerăm ADN-ul ca o abordare evolutivă nouă - ca o modificare a ARN-ului, specializată în scopul îndeplinirii funcției și salvării de copii unice ale genelor în depozitul genomului clitinei fără participarea indirectă la biosinteza proteinelor. .
În plus, de îndată ce a fost utilizat ARN-ul activ catalitic, ideea de primație a ARN-ului în viață a avut un efect puternic spre dezvoltare, iar conceptul a fost formulat. lumea autosuficientă a ARN-ului, scho suflă viața actuală [ , ]. O posibilă schemă a ARN viknennia este prezentată în fig. 6.
Sinteza abiogenă a ribonucleotidelor în aceeași asociere covalentă în oligomerul și polimerul de tip ARN a putut fi observată aproximativ în aceleași minți și în aceleași condiții chimice care au fost postulate pentru sinteza aminoacizilor și polipeptidelor. Recent, O.B. Chetverin și colegii săi (Institute of Protein RAS) au arătat experimental că poliribonucleotidele (ARN) sunt utilizate în mediul acvatic primar la recombinare spontană, astfel încât schimbul de lanceolate lanceolate prin calea de transesterificare ї. Schimbul de coaste scurte lancete pentru o lungă perioadă de timp este responsabil pentru reducerea poliribonucleotidelor (ARN), iar recombinarea în sine este similară cu diversitatea structurală a acestor molecule. Printre acestea, moleculele de ARN catalitic active ar putea fi învinuite.
A produce apariția unor molecule de ARN unice, care ar putea cataliza polimerizarea ribonucleotidelor sau splicing-ul oligonucleotidelor pe lancea complementară ca și pe matrice [ ], a însemnat formarea mecanismului de replicare a ARN. Replicarea catalizatorilor de ARN (ribozime) înșiși are o mică consecință în cauzarea populațiilor de ARN auto-replicabile. Producerea propriilor copii de ARN sa multiplicat. Iertări inevitabile în copiere (mutație) și recombinare în populațiile de ARN, care se auto-replica, au creat din ce în ce mai multă diversitate în întreaga lume. În acest rang, transferul lumii antice a ARN - ce „o lume biologică autosuficientă, în care moleculele de ARN au funcționat ca material genetic și ca catalizatori asemănătoare enzimelor” .
Justificarea biosintezei proteinelor. În plus, pe baza lumii ARN-ului, există puține dovezi ale formării mecanismelor pentru biosinteza proteinelor, apariția diferitelor proteine dintr-o structură și dominanță degradate, compartimentarea sistemelor pentru biosinteza proteinelor și seturi de proteine, poate sub formă de coacervate și evoluții Ultima structură rămasă a clitinei este clitina vie (div. Fig. 6) . ).
Problema tranziției de la lumea antică a ARN-ului la lumea actuală de sinteză a proteinelor este importantă pentru o soluție fundamental teoretică. Posibilitatea sintezei abiogene a polipeptidelor și a discursurilor asemănătoare proteinelor nu ajută la rezolvarea problemei, nu există semne ale vreunei căi specifice, ca sinteza straturilor mig bibuty din ARN și sub control genetic. Controlul genetic al sintezei polipeptidelor și proteinelor se dezvoltă independent de sinteza abiogenă primară, în felul său, cu îmbunătățirea ARN-ului deja existent. În literatura de specialitate, au fost propuse câteva ipoteze pentru mecanismul modern de biosinteză a proteinelor în lumea ARN, dar, poate, ele pot fi văzute ca gândite în detaliu și fără o privire asupra capacităților fizice și chimice. Depun versiunea mea a procesului de evoluție și specializare a ARN, care a dus la sfârșitul aparatului de biosinteză a proteinelor (mic 7), dar nu pretind că am terminat.
S-a propus o schemă ipotetică pentru a răzbuna două sute de momente, care par a fi importante.
Primul, Se postulează că oligoribonucleotidele sintetizate abiogen au fost recombinate activ în spatele mecanismului suplimentar de transesterificare spontană neenzimatică, ducând la reducerea lancetelor de ARN și dând naștere diversității acestora. În același mod, în populația de oligonucleotide și polinucleotide ar putea apărea atât specii de ARN (ribozime) active catalitic, cât și alte specii de ARN cu funcții specializate (Fig. 7). Mai mult decât atât, recombinarea neenzimatică a oligonucleotidelor care se leagă în mod complementar la matrița polinucleotidică ar putea asigura fuziunea (splicing) fragmentelor complementare acestui șablon într-o singură lance. În acest fel, în loc să catalizeze polimerizarea mononucleotidelor, s-ar putea efectua copiarea (multiplicarea) primară a ARN-ului. Presupunând că erau ribozime care aveau o activitate redusă a polimerazei, atunci eficiența (acuratețea, flexibilitatea și productivitatea) copierii pe complementare. matricele sunt mici și cresc semnificativ.

Mal. 7. Schema de evoluție și specializare a moleculelor de ARN
în procesul de trecere de la lumea antică a ARN-ului la la lumea actuală
biosinteza proteinelor determinată genetic
Alte Un punct important în versiunea mea este că primul aparat pentru biosinteza proteinelor a apărut în eradicarea multor specii de ARN specializat înainte de apariția aparatului pentru replicarea enzimatică (polimeraza) a materialului genetic - ARN și ADN. Aparatul lor primar, incluzând ARN proribozomal activ catalitic, are activitate peptidil transferază scăzută; recrutarea de pro-ARNt care leagă în mod specific aminoacizii sau peptidele scurte; alt ARN proribozomal, interacționând simultan cu ARN proribozomal catalitic, pro-ARNm și pro-ARNt (div. Fig. 7). Un astfel de sistem ar putea deja sintetiza polipeptide lanceolate sub forma unei reacții de transpeptidare catalizată. Printre celelalte proteine active catalitic - enzimele primare (enzimele) - au existat și proteine care catalizează polimerizarea nucleotidelor - replici, sau polimeraze NK.
Vtim, poate, despre ce este ipoteza lumină străveche ARN, ca lider al lumii vii de astăzi, nu poate fi suficient pentru a fundamenta problema principală - o descriere științific plauzibilă a mecanismului de tranziție de la ARN și replicare la biosinteza proteinelor. Ipoteza alternativă a lui A.D. Altshtein (Institutul de Biologie Genetică, Academia Rusă de Științe), se postulează că replicarea materialului genetic și traducerea iogo - sinteza proteinelor - vinicals și a evoluat o oră la un moment dat, pornind de la interdependența sintetizării abiogene a oligonucleotidelor în i aminoacil- nucleotidilaţii - anhidride mixte. Vai, deja vine cazacul... ( „Eu Shahrazad am prins rana și ea a fixat permisiunea de a promova”.)
Literatură
1. Watson JD, Crick F.H.C. Structura moleculară a acizilor nucleici // Natura. 1953. V. 171. P. 738-740. 2. Watson JD, Crick F.H.C. Implicații genetice ale structurii acizilor nucleici dezoxiriboze // Nature 1953 V. 171. P. 964-967. 3. Spirin A.S. Biologie modernă și siguranță biologică // Buletinul Academiei Ruse de Științe. 1997. Nr. 7. 4. Spirin A.S. Despre structura macromoleculară a chimiei ribonucleare naturale cu polimeri înalți // Journal of Molecular Biology. 1960. V. 2. P. 436-446. 5. Kirn S.H., Suddath F.L., Quigley GJ. că în. Structura terțiară tridimensională a ARN-ului de transfer al fenilalaninei de drojdie // Știință. 1974. V. 185. P. 435-40. 6. Robertas J.D., Ladner J.E., Finch J.T. că în. Structura ARNt-ului fenilalaninei de drojdie la rezoluție 3 A // Natura. 1974. V. 250. P. 546-551. 7. Vasiliev V.D., Serdyuk I.N., Gudkov A.T., SPIRin A.S. Autoorganizarea ARN-ului ribozomal // Structura, Funcția și Genetica Ribozomilor / Eds. Hardesty B. şi Kramer G. New York: Springer-Verlag, 1986, p. 129-142. 8. Baserga SJ., Steitz J.A. Raznomanіtny svіt malikh ribo-nucleoproteine // The ARN World / Eds. Gesteland R.F. și Atkins J.F. New York: Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1993, pp. 359-381. 9. Kruger K., Grabowski PJ., Zaug AJ. că în. ARN auto-splicing: autoexcizia și autociclizarea secvenței de intervenție a ARN-ului ribozomal a Tetrahimena// celulă. 1982. V. 31. P. 147-157. 10. Guerrier-Takada C., Gardiner K., Marsh T. et al. Ribonucleazele P este o subunitate catalitică a enzimei // Celulă. 1983. V. 35. P. 849-857. 11. Oparin A.I. Viața de plimbare. M: Moscova Robotnik, 1924. 12. Oparin A.I. Viața Vinnyknennya a Pământului (al treilea fel.). M: Vedere a Academiei de Științe a SRSR, 1957. 13. vai S. Evoluția codului genetic // Codul genetic. New York: Harper & Row, 1967, pp. 179-195. 14. Crick F.H.C. originea codului genetic // Journal of Molecular Biology. 1968. V. 38. P. 367-379. 15. Orgel L.E. Evoluția aparatului genetic // Journal of Molecular Biology. 1968. V. 38. P. 381-393. 16. Gilbert W. Lumea ARN // Natura. 1986. V 319 p. 618. 17. Joyce G.F., Orgel L.E. Perspective pentru înțelegerea originii lumii ARN // The ARN World / Eds. Gesteland R.F. și Atkins J.F. New York: Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1993 P 1-25. 18. Chetverina H.V., Demidenko A.A., Ugarov V.I., Chetverin A.B. Reorganizare spontană în secvențe de ARN // Scrisori FEBS. 1999. V. 450. P. 89-94. 19. Bartel D.P., Szostak J.W. Izolarea noilor ribozime dintr-un grup mare de secvențe aleatorii // Știință. 1993. V. 261. P. 1411-1418. 20. Ekland E.H., Bartel D.P. Polimerizarea ARN-ului catalizat de ARN cu trifosfați nucleozidici // Nature. 1996 V. 382. P. 373-376. 21. Orgel L.E. originea vieții - Trecere în revistă a faptelor și speculațiilor // Trends in Biochemical Sciences. 1998. V. 23. str. 491-495. 22. Altstein A.D. Sistemul genetic Pohodzhennya: ipoteza descendenței // Biologie moleculară. 1987. T. 21. S. 309-322.








