Em todas as células vivas, as proteínas são sintetizadas pelos ribossomos. . O ribossomo é uma grande macromolécula com uma estrutura assimétrica dobrada em um quarto, inspirada em ácidos ribonucleicos (RNA ribossômico) e proteínas. Para sintetizar proteínas, o ribossomo deve ser protegido:
1. Um programa que define a ordem de coleta de resíduos de aminoácidos na lança polipeptídica da proteína.
2. Material de aminoácidos, do qual será utilizada a proteína.
3. Energia.
O próprio ribossomo tem função catalítica (enzimática), responsável pela incorporação de ligações peptídicas e, aparentemente, polimerização de resíduos de aminoácidos na lança polipeptídica da proteína.
O programa, que define a ordem dos resíduos de aminoácidos na lança polipeptídica da proteína, surge na forma de ácido desoxirribonucléico (DNA), ou seja, do genoma da clitina. A síntese das lanças de RNA é complementar a uma das lanças de DNA i, de modo que confirma exatamente a sequência desoxirribonucleotídica da outra lança de DNA em sua sequência de ribonucleotídeos. O processo dessa cópia de um gene, que é modificado pela enzima RNA polimerase, retira o nome da transcrição. O RNA durante a síntese deste último, especialmente em clitinas eucarióticas, pode estar sujeito a uma série de alterações adicionais, chamadas de processamento, no decorrer das quais podem ser transformadas em pequenas partes da sequência de nucleotídeos. O RNA, que entra, entra nos ribossomos como um programa, que determina a sequência de aminoácidos na proteína sintetizada. É chamado de RNA informativo ou "mensageiro" (mRNA). Dessa forma, a própria transcrição dos genes e a adoção do mRNA garantem o fluxo de informações do DNA para os ribossomos.
O material da espiga, do qual a proteína será, é aminoácidos. No entanto, os aminoácidos não são vicários pelo ribossomo. Para servir de substrato para o ribossomo, o aminoácido deve ser ativado para a participação na divisão resultante do ATP e é aceito (ligado covalentemente) por uma molécula especial de RNA, chamado de transferência ou RNA de transferência (tRNA), com ajuda adicional. Otrimani aminoacil-tRNA está presente no ribossomo como um substrato para a síntese de proteínas. Além disso, a energia da ligação química entre o excesso de aminoácidos e o tRNA é vicorada pela reação da ligação peptídica no ribossomo. Assim, a ativação de aminoácidos e a adoção de aminoacil-tRNA fornecem material e energia para a síntese de proteínas ribossômicas.
Os três fluxos (informação, material e energia) estão agrupados em ribossomos. Ao aceitá-lo, o ribossomo transfere, ou traduz, a informação genética da sequência de nucleotídeos do mRNA para a sequência da sequência de aminoácidos da proteína lanceta polipeptídica sintetizada. Yakshcho Vyavita Ts em termos moleculares, então o ribossoma do subganum de Lantsyug MRNA (correr para o naufrágio de não-escolas), fui ligado ao soressine da tribo aminoacil. mRNA dividido. Além disso, o problema do código genético é o culpado: quais combinações de nucleotídeos determinam, ou seja, qual código a pele com 20 aminoácidos, a partir de quais moléculas de proteína serão formadas?
A rotação do ribossomo do mRNA da lanceta (ou, em outras palavras, a passagem da lanceta do mRNA pelo ribossomo) estabelece uma estrita ordem timcásica de entrada no ribossomo de vários aminoacil-tRNAs, dependendo da ordem dos expansão das combinações de nucleotídeos que codificam, reduzem o mRNA. O excesso de aminoacil do aminoacil-tRNA reverso é ligado covalentemente pelo ribossomo à lança polipeptídica em crescimento. O tRNA desacilado é liberado dos ribossomos nos locais. Assim, sequencialmente, passo a passo, haverá uma lança polipeptídica da proteína (div. esquema 1).
Biossíntese de proteínas.
Troca de plástico (assimilação e anabolismo) - a sequência de reações de síntese biológica. O nome desse tipo de troca é dia a dia: a partir das falas, como a postura de um clitin, se estabelecem falas, semelhantes às falas de um clitin.
Vejamos uma das formas mais importantes de troca de plástico - a biossíntese de proteínas. Biossíntese de proteínas encontrado em todos os pró-eucariotos de clitina. As informações sobre a estrutura primária (ordem dos aminoácidos) de uma molécula de proteína são codificadas pela sequência de nucleotídeos na divisão da molécula de DNA - gene.
O gene de uma molécula de DNA, que determina a ordem dos aminoácidos em uma molécula de proteína. Além disso, na forma de nucleotídeos no gene, a ordem dos aminoácidos no polipeptídeo deve ser determinada. Esta é a estrutura primária, em vista da qual todas as outras estruturas, o poder e a função da molécula de proteína, encontram-se em sua própria linha.
O sistema de registro da informação genética no DNA (i RNA) na sequência aparente de nucleotídeos é chamado de código genético. Tobto. unidade de código genético (códon) - um trio de nucleotídeos no DNA ou RNA que codifica um aminoácido.
Nosso código genético inclui 64 códons, dos quais 61 são codificantes e 3 não codificantes (códons terminadores, que indicam a finalização do processo de tradução).
Terminadores de códons i - RNA: UAA, UAG, UGA, no DNA: ATT, ATC, ACT.
O processo de tradução é iniciado pelo iniciador de códon (AUG, DNA - TAC), que codifica o aminoácido metionina. O códon cei é o primeiro a entrar no ribossomo. A propósito, a metionina, embora não seja transferível, como o primeiro aminoácido dessa proteína, é usada.
O código genético pode ser característico do poder.
1. Universalidade - o código do mesmo para todos os organismos. Um e o mesmo tripleto (códon) em qualquer organismo codifica o mesmo aminoácido.
2. Especificidade - o códon da pele codifica apenas um aminoácido.
3. Virogenidade - mais aminoácidos podem ser codificados pelo número de códons. Existem 2 aminoácidos - metionina e triptofano, que só podem ser usados para uma variante do códon.
4. Genes Mіzh є “sinais diferentes” - três trigêmeos especiais (UAA, UAG, UGA), cujas peles indicam a síntese do polipeptídeo lanciug.
5. Não há meios do gene de "signos diferentes".
Para sintetizar a proteína, informações sobre a sequência de nucleotídeos em sua estrutura primária podem ser entregues aos ribossomos. Este processo inclui duas etapas - transcrição e tradução.
Transcrição(reescrita) a informação é obtida por meio de síntese em uma das lanças da molécula de DNA da molécula de RNA de lanceta única, a sequência de nucleotídeos é exatamente a mesma que a sequência de nucleotídeos na matriz - a lança polinucleotídica do DNA .
Vaughn (i - RNA) é um intermediário que transmite informações do DNA para o local de seleção de moléculas de proteína nos ribossomos. A síntese de i RNA (transcrição) é considerada uma classificação ofensiva. A enzima (RNA - polimerase) divide a lanceta sublingual do DNA, e uma das lancetas (codificação) segue o princípio da complementaridade, os nucleotídeos do RNA são vibrados. A molécula de RNA é sintetizada dessa maneira (síntese de matriz) e entra no citoplasma, e pequenas subunidades de ribossomos são enfileiradas em uma extremidade.
Outra etapa na biossíntese de proteínas é transmissão- tse tradução da sequência de nucleotídeos na molécula e - RNA na sequência de aminoácidos no polipeptídeo. Em procariotos, que não podem formar um núcleo bem formado, os ribossomos podem se ligar a uma molécula de i-RNA recém-criada imediatamente após a divisão do DNA ou acumular até a conclusão completa da síntese. Em eucariotos, o i-RNA pode ser transportado para a teia de aranha através da membrana nuclear para o citoplasma. A transferência é causada por proteínas especiais, que formam um complexo com uma molécula de RNA. As funções de creme de transferência de proteínas citoplasmáticas são protegidas por i - RNA de diy, que permite enzimas citoplasmáticas.
No citoplasma, uma das pontas do i - RNA (e aquela que inicia a síntese de uma molécula no núcleo) entra no ribossomo e inicia a síntese do polipeptídeo. No mundo, passando pela molécula de RNA, o ribossomo traduz tripleto após tripleto, adicionando sucessivamente aminoácidos à extremidade crescente da lanceta polipeptídica. A correspondência exata do aminoácido com o código trigêmeo e o RNA é assegurada pelo t RNA.
O RNA de transporte (t-RNA) "traz" aminoácidos para a grande subunidade do ribossomo. A molécula de t-RNA pode ter configuração dobrada. Em alguns locais entre nucleotídeos complementares, ligações de água são estabelecidas e a molécula tem a forma de uma folha estável. No topo das dobras, há um trio de nucleotídeos livres (anticódon), que contém um único aminoácido, e a base contém uma cadeia de aminoácidos (Fig. 1).
Mal. 1. Esquema de transporte RNA: 1 - links de água; 2 - anticódon; 3-misce aminoácidos anexados.
O t-RNA da pele só pode transferir seu próprio aminoácido. O T-RNA é ativado por enzimas especiais, adicionando seu próprio aminoácido e transportando-o para o ribossomo. No meio dos ribossomos no momento da pele existem apenas dois códons de i-RNA. Uma vez que o anticódon do t-RNA é complementar ao códon do i-RNA, há um advento oportuno do t-RNA com um aminoácido para o i-RNA. Antes do próximo códon vem outro t-RNA, como se carregasse seu aminoácido. Os aminoácidos são misturados com a ajuda da grande subunidade do ribossomo e, com a ajuda de enzimas, as ligações peptídicas são estabelecidas com eles. A ligação entre o primeiro aminoácido e o її t-RNA, e o t-RNA vai do ribossomo atrás do aminoácido atacante. O ribossomo move um trigêmeo e o processo é repetido. Dessa forma, a molécula do polipeptídeo é construída de forma incremental, na qual os aminoácidos são misturados em uma sequência de sequências na ordem dos tripletos que os codificam (síntese de matriz) (Fig. 2).

Mal. 2. Esquema para síntese de proteínas: 1 – i-RNA; 2 - subunidades do ribossomo; 3 – t-RNA com aminoácidos; 4 – t-RNA sem aminoácidos; 5 - polipeptídeo; 6 - códon ta-RNA; 7-anticódon do tRNA.
Um ribossomo é capaz de sintetizar uma nova lança polipeptídica. Prote, muitas vezes uma molécula de i-RNA colapsa uma espadilha de ribossomos. Esses complexos são chamados de polirribossomos. Após a conclusão da síntese, as lanças polipeptídicas são fundidas na forma de uma matriz - moléculas de i-RNA, se enrolam em uma hélice e ganham uma poderosa estrutura (secundária, terciária ou quartel). Os ribossomos funcionam ainda com mais eficiência: ao esticar 1c, o ribossomo bacteriano converte a lanceta polipeptídica de 20 aminoácidos.
A biossíntese de proteínas (polipeptídeos) é um enovelamento supra-adventício e um processo maravilhoso. A biossíntese de proteínas ocorre ativamente em todos os órgãos e tecidos, incluindo eritrócitos. Muita clitina sintetiza proteínas para "exportar" (clitina hepática, folículos subescapulares), e assim o mau cheiro vai vingar até mesmo um grande número de ribossomos. Nas células animais, o número de ribossomos chega a 10 5, o diâmetro do ribossomo chega a 20 nm.
O processo de síntese de proteínas ocorre no meio das células na superfície dos ribossomos, como complexos de duas subunidades com constante de sedimentação de 60S e 40S, que funcionam como uma única unidade. Nos ribossomos, as proteínas tornam-se 30-35% e o RNA ribossômico - 65-70%. Os ribossomos têm fendas de aminoacil e peptidil. A primeira serve para a fixação do complexo de aminoácidos ativos e tRNA, que se localiza no ribossomo, e a outra fixa a lança polipeptídica, ligando-se a outro tRNA. As subunidades dos ribossomos são sintetizadas no núcleo do núcleo em um molde de DNA.
A essência do processo de síntese de proteínas é representada pelo esquema:
O sistema de síntese de proteínas inclui ribossomos, ácidos nucléicos, um conjunto de 20 aminoácidos, várias enzimas, ATP, GTP, íons de magnésio e cerca de 200 fatores proteicos não catalíticos diferentes.
A molécula de proteína é uma longa linha de resíduos de aminoácidos, que está presente na forma média de 100 a 500 aminoácidos. O programa para a síntese de proteínas da pele é armazenado na molécula de ácido desoxirribonucléico (DNA). A molécula de DNA é um polímero, cujos monômeros são nucleotídeos. A sequência de bases nitrogenadas em uma molécula de DNA determina a sequência de aminoácidos em uma molécula de proteína.
A molécula de DNA possui várias bases nitrogenadas: adenina (A), guanina (G), citosina (C) e timina (T). Uma sequência de três subestações (tripleto) para se tornar um códon, que requer um único aminoácido.
Ácidos nucleicos - DNA e RNA - componentes obrigatórios da biossíntese de proteínas. O DNA é responsável pela conservação da informação genética, enquanto o RNA é responsável pela transferência dessa informação e sua implementação no aparecimento de moléculas de proteína. Pode-se argumentar que a principal função do DNA é preservar o genótipo e o RNA - para o genótipo do vírus.
No plano do cliten, prevalece o RNA ribossômico (rRNA). rRNA pode espiralizar a plaqueta, impedir a modificação de nucleotídeos (por exemplo, 2-metil-ribose). O rRNA torna-se próximo a 80% do RNA total na clitina. Outro tipo de RNA em representações clitíneas de RNA de transporte (tRNA), iaque, iaque e outros tipos de RNA, é sintetizado no núcleo. 10-15% da quantidade total de RNA na clitina cai na parte її. Mais de 60 tRNAs diferentes foram revelados. Além disso, o transporte de outros aminoácidos é baseado em um pequeno número de diferentes tRNAs. Para os aminoácidos da pele, as clitinas aceitam um tRNA específico. As moléculas de tRNA são divididas aleatoriamente. Suas estruturas têm 75-93 ribonucleídeos.
O aminoácido se liga ao grupo 3-OH livre do mononucleotídeo terminal do tRNA, representado pelo ácido adenílico. O tRNA é a parte mais importante - um anticódon, para o qual o complexo de aminoácidos e tRNA é determinado pela sequência de três nucleotídeos no RNA da matriz (códon). O anticódon e o códon buscam complementarmente ajuda para conexões de água.
Embora carregue informações de decaimento no clitino e no DNA, por estar centralizado no núcleo, e se a síntese de proteínas for realizada no citoplasma, então, também, pode haver um mediador que transmita informações ao citoplasma do clitino. O mediador cym foi o RNA mensageiro informativo (mRNA). O mRNA é precedido por 2% da quantidade total de RNA de clitina. Moléculas de mRNA encontradas (incluem até 5 mil nucleotídeos). O mRNA também contém bases nitrogenadas. Três deles (A, G, C) são os mesmos do DNA e o quarto é o uracil.
A informação codificada no mRNA é necessária para a síntese de uma molécula de proteína que se encontra nos ribossomos. A síntese de mRNA no núcleo das células é bastante lenta, o que é necessário para a biossíntese ativa de moléculas de proteína. O mRNA se estabelece em uma das fitas de DNA no núcleo. Nesse caso, a estrutura de fita dupla do DNA se desenrola e, com a participação da RNA polimerase depletada de DNA, a síntese do mRNA segue o princípio da complementaridade:

Esquema para síntese de mRNA
O princípio da complementaridade significa que a adenina na hélice do DNA é combinada com o mRNA do uracil, a timina com a adenina e a guanina com a citosina. Além disso, o mRNA lê as informações do DNA.
A etapa DNA - RNA, dessa forma, significa a síntese da molécula de mRNA, na qual a sequência de nucleotídeos é complementar à divisão do canto (gene) do DNA. Este processo é chamado de transcrição. Em seguida, o mRNA está localizado no ribossomo, um a um com її subunidades. Uma molécula de mRNA é fixada em vários ribossomos ao mesmo tempo, formando os chamados polissomos. A presença de polissomos promove a eficiência e flexibilidade da vicariação do mRNA.
A síntese da lanceta polipeptídica do armazém variante é realizada na matriz de mRNA. O processo de transferência de mRNA para uma proteína omitindo o nome da tradução. A etapa “RNA -> proteína” é o processo de síntese de proteínas, que direciona o mRNA. Desta forma, a transferência de informação zavzhdi ir direto DNA - RNA - proteína.
O processo de tradução inclui as seguintes etapas:
A mesma enzima participa da fixação de um aminoácido ativado para frente na posição 2 ou 3 da ribose do nucleotídeo restante do tRNA:
Nesse complexo de aminoácidos, o ribossomo é transportado e ocorre a síntese de uma molécula de proteína. A aminoacil-tRNA sintetase é específica, pode reconhecer como um aminoácido, i-tRNA. Na clitina, nesta ordem, existem pelo menos 20 sintetases diferentes, variando até o número de a-aminoácidos.
2. tRNA, ligado por uma ligação etérea com um único aminoácido, localiza-se no ribossomo e inter-relaciona-se com o mRNA para o tipo de complementaridade entre um trio específico de nucleotídeos no mRNA, o nome códon, e o trio específico complementar de nucleotídeos no mRNA (anticódon). Desta forma, o códon da pele do mRNA fornece fixação específica de um aminoácido na lança peptídica para a ajuda do anticódon do tRNA. O ribossomo passa sobre as moléculas de mRNA, lendo sequencialmente todos os códons, estabelecendo assim a ordem de dissolução de todos os aminoácidos que são entregues ao sítio de síntese.
A síntese da molécula de proteína vai diretamente do grupo amino livre para o grupo carboxila livre do aminoácido. O nome do aminoácido na espiga na síntese do polipeptídeo lanciug é metionina, cujo códon é a sequência de nucleotídeos do mRNA AUG.
A iniciação da síntese de polipeptídeos começa com a fixação de dois anticódons de tRNA atrás dos segundos códons de mRNA. O processo de aumentar a visibilidade da energia, que serve como GTP, bem como a participação de uma série de fatores proteicos iniciadores e peptidiltransferase.
Para a participação desta enzima, a velocidade de iluminação ligações covalentes atingir 1200 aminoácidos / xv / ribossomos.

Esquema de iniciação para a síntese de polipeptídeos
3. Depois que o dipeptídeo “indesejado” é introduzido, o tRNA preenche o ribossomo e pode fornecer novas moléculas de aminoácidos, e o mRNA é empurrado através dos ribossomos (polissomos) por três nucleotídeos. Como resultado da relocação (translocação), o códon livre assume uma posição de reconhecimento da molécula de tRNA central. Além disso, na fase de alongamento, ocorre a sequência de adição de um aminoácido à lanceta polipeptídica na ordem dos códons da molécula de mRNA.
A lança polipeptídica, que supostamente está ligada a uma molécula de tRNA, é fixada na subunidade maior do ribossomo. A adição do aminoácido suplementar dérmico à lança polipeptídica se deve à relação entre o grupo amino do aminoácido, que se une no complexo com o tRNA e o grupo carboxila do peptídeo.
4. Término ou finalização da síntese da molécula polipeptídica pela obtenção da terminação códon "sem sentido" e fator de terminação proteica. Existem três códons (UAH, UGA, UAA), que não codificam, não se ligam a um aminoácido, pois as células não possuem anticódons de tRNAs complementares a elas. Teoricamente, apenas um códon "sem sentido", que é o responsável pela síntese da molécula de proteína, é responsável pela passagem do mRNA 5-3 pelo polissomo.
A presença de um códon de terminação em qualquer divisão do mRNA significa terminação síntese proteíca. Como resultado, o polissomo se desintegra, o mRNA do nevicoristano é hidrolisado pela polinucleosídeo fosforilase e as subunidades do ribossomo são preparadas para iniciar a síntese de uma nova molécula de proteína.
O mRNA pode participar repetidamente do processo de biossíntese de proteínas. A trivalência do funcionamento da molécula de mRNA não é a mesma em diferentes organismos. Vaughn pode oscilar de alguns assobios a alguns dib.
5. No DNA, a estrutura primária da proteína é codificada. Portanto, a síntese nos ribossomos da molécula de proteína ainda pode ser um estado residualmente concluído. Eles representam os polipeptídeos primários, que então reconhecem modificações numéricas (associação de monômeros com oligômeros estabelecidos, adição de coenzimas, transformações químicas) que alteram a estrutura das proteínas e, portanto, sua atividade.
As estruturas secundárias e terciárias não são codificadas, elas são determinadas pelo poder da estrutura primária, e isso significa que essa forma chi insha da molécula de proteína é depositada na sequência de aminoácidos e as possibilidades de suas interações entre si. As modificações estruturais das proteínas, que são sintetizadas, podem ser mais comuns em ribossomos iguais ou após a conclusão da síntese como resultado da adição de outros grupos funcionais.
Olhou para o esquema de transmissão à vista
você pode mudar de maneiras diferentes. Assim, nos vírus, que não vingam o DNA, a informação está embutida no RNA. Quando o vírus penetra na clitina, a informação é transferida para o DNA da clitina, e o restante já está sintetizando o mRNA, no qual as proteínas do vírus são sintetizadas na matriz. Esse processo é chamado de transcrição reversa, e o esquema de transmissão de qualquer forma será ofensivo:
Enquanto a sequência de nucleotídeos no DNA e, portanto, no mRNA for preservada, a natureza da proteína recém-sintetizada permanecerá inalterada.
A informação genética necessária para a síntese da proteína pode ser apresentada de forma semelhante ao registro da linguagem humana, pois é formada a partir da sequência de letras que formam as palavras dessa fala. Na linguagem genética, entretanto, há menos letras chotiri - bases chotiri (adenina, guanina, uracil, citosina).
O código genético inclui palavras triliterais. Chotiri sugere desta forma (43) dar 64 opções (palavras), que são mais ou menos suficientes, para codificar 20 aminoácidos. Nesta ordem, 64 códons compõem o código genético (Tabela 3).
A análise do código genético mostra que o número de códons é diferente para diferentes aminoácidos. Por exemplo, metionina e triptofano podem ter apenas um códon, enquanto arginina, leucina e serina podem ter seis códons cada. A presença de vários códons para um aminoácido reflete a "virogeneidade" do código. Além disso, o próprio aminoácido pode ser codificado como um kilkom para seus próprios trigêmeos de nucleotídeos. Ao mesmo tempo, o trigêmeo da pele contém um aminoácido inteiro na lanceta polipeptídica, que é sintetizada.
Tabela 3
Código genético
|
nucleotídeo |
Outro nucleotídeo |
nucleotídeo |
|||
O código genético é universal e o mesmo em espécies de diferentes desenvolvimentos (humanos, criaturas, plantas, microorganismos). A universalidade do código é dizer que todos os organismos vivos do passado são um pequeno ancestral.
Os aminoácidos Okremi (hidroxiprolina, oxilisina), por exemplo, não mexem com o códon e se contentam com ajuda reações químicas já após a síntese do polipeptídeo lanceug. Este processo, tendo tomado o nome de modificação pós-traducional e ainda mais importante para o bom funcionamento da proteína da pele.
Códons silenciosos (UAA, UAG, UGA) não codificam aminoácidos, a proteína é realmente um sinal para a conclusão da síntese de uma molécula de proteína.
Assim, o mRNA é um transportador ininterrupto de informação genética do núcleo para o ribossomo do citoplasma. Um ribossomo empresta aproximadamente 80 nucleotídeos por mRNA e catalisa aproximadamente 100 ligações peptídicas por quilina (Severin E. S. et al., 2011).
A síntese de moléculas de proteína pode estar sujeita a modificações estruturais nos ribossomos iguais ou após a conclusão da síntese como resultado da adição de diferentes grupos funcionais. O mRNA citoplasmático pode ter um período relativamente curto. A quantidade de mRNA é sintetizada e armazenada na forma inativa, estando pronta para rápida síntese proteica. As informações do mRNA de Oscilki estão relacionadas à sequência linear de nucleotídeos, a integridade da sequência é extremamente importante. Se uma alteração na ordem dos nucleotídeos pode ou não alterar a síntese da proteína. Um baixo número de inibidores da replicação do DNA em organismos clitínicos (antibióticos, lontras químicas, drogas antivirais) foi estabelecido este ano. Mudanças na sequência de bases purínicas ou pirimídicas no gene retiraram o nome da mutação.
A substituição de mais de um nucleotídeo em um códon (mutação) leva a uma alteração na codificação de um aminoácido por outro. Por exemplo, uma mutação devido à substituição do ácido glutâmico por valina na molécula de hemoglobina leva à síntese de hemoglobina, que leva à anemia falciforme. Hoje, mais de 200 mutações do polipeptídeo cola da molécula de hemoglobina humana foram relatadas. Muitas vezes são mutagênicos da fala (nitrosaminas, por exemplo), que alteram a estrutura das bases nitrogenadas, o que leva a uma mudança na natureza da complementaridade das bases. A reflexão ultravioleta causou a condensação do excesso de timina com os dímeros de timina dissolvidos. Felizmente, após o forte influxo de mudanças ultravioleta, as criaturas são protegidas pelo balão de ozônio da atmosfera.
Muitos antibióticos encontrados na prática veterinária inibem a síntese de proteínas bacterianas (lincomicina, eritromicina, cloranfenicol) no estágio de tradução. Com esta clitina microbial gyne chi cresce o seu desenvolvimento. Esses antibióticos, como as tetraciclinas, não interferem na síntese ribossômica em seres vivos. As penicilinas não são inibidores diretos da síntese proteica, atuam como inibidores de bactérias ao bloquear a síntese de hexapeptídeos na parede celular. Deve-se notar que a síntese de proteínas ocorre não apenas nos ribossomos, mas também nas mitocôndrias. As mitocôndrias podem ter um aparato independente de síntese de proteínas para suas necessidades, embora nem todas as proteínas mitocondriais sejam sintetizadas nessas organelas. O RNA mitocondrial torna-se menos de 3% da quantidade de RNA da clitina. Os ribossomos das mitocôndrias são menores atrás das bordas, mais baixos são citoplasmáticos. O códon UGA, como terminador da síntese de proteínas no citoplasma, é encontrado nas mitocôndrias próximo ao códon UGG para codificação de aminoácidos.
A síntese de proteínas nos ribossomos não cria um corpo residualmente completo. Eles representam os polipeptídeos primários, que então reconhecem modificações numéricas (associação de monômeros com oligômeros, adição de coenzimas, transformações químicas) que modificam a estrutura da proteína e, portanto, sua atividade.
Figura 9 da apresentação “Biossíntese de proteínas”às aulas de biologia sobre o tema "Biossíntese de proteínas"
Tamanho: 960 x 720 pixels, formato: jpg. Para baixar gratuitamente a imagem da aula de biologia, clique na imagem com o botão direito do mouse e clique em "Salvar imagem como...". Para mostrar as fotos da aula, você também pode baixar a apresentação "Biosynthesis of protein.pptx" com todas as fotos em arquivo zip. Arquivo Rozmir - 1719 KB.
Fazer uma apresentação“Funções dos brancos” - É assim que são percebidas a recepção de sinais do ambiente externo e a transferência de informações para o cliente. Com a quebra de 1 g de proteína em produtos finais, observa-se 17,6 kJ. O que é renaturação? Podіb'єmo pіdbags: 9. Catalítico. O processo de restaurar a estrutura da proteína após a desnaturação é chamado de renaturação. Pimenov A.V. As proteínas são uma das fontes de energia das células.
"Proteínas da fala" - Por exemplo: colágeno. Biologia de Vikladach: Boldireva L.A. . Bunda: ovo cozido. Aminoácido - discurso orgânico, Proteínas não resolvidas - fibrilares. Brancos de Kharchov. . Proteínas Zahisnі. Estrutura da proteína. Organismo Vikoristovuyutsya para ruhu. Proteínas energéticas.
"Proteínas e funções do yoga" - Papel catalisador. Função do motor. Conceito de proteína. A hidrólise de proteínas leva à clivagem de ligações polipeptídicas: Visnovok: Vasos portadores de sangue, tendões e cabelos foram induzidos a partir de proteínas. Budova e funções da proteína. poder químico bilkiv. as proteínas participam das membranas estabelecidas de clitina, organoides e membranas de clitina.
"Biossíntese de proteínas" - Lista de literatura. Introdução 4. Zmist. Biossíntese de proteínas em células vivas. 7. 10. 9. Esquema do clitin crescente e vivo. 5. 6. 1. 8. 2. 3.
"Biossíntese de proteínas" - Tradução (lat. Transferência, tradução). Transcrição (lat. reescrita). Inverta-se. Significado de branco. Zmist. Biossíntese de energia. O papel das enzimas. Síntese do polipeptídeo lanciug em ribossomos. 5. Qual é a sequência de nucleotídeos no i-RNA registrada no cruzamento de DNA: T-A-C-G-G-A-T-C-A-C-G-A -G-T-G-C-T A-U -G-Ts-G-U-A-G-U-G-Ts-U A-U-G-Ts-Ts-U-A-G-U -G-Ts-U.
"Biossíntese da biologia de proteínas" - Mikola Kostyantinovich Koltsov (1872-1940). AG A principal função dos ribossomos é a síntese de proteínas. O dogma central (postulado básico) da biologia molecular é a síntese de matrizes. C. Anticódon - um trio de nucleotídeos no topo do tRNA. Biossíntese de proteínas. Após a conclusão da síntese, o iRNA se decompõe em nucleotídeos.
Os temas têm 8 apresentações no total
© A. S. espirina
BIOSINTEZ BILKIV, SVIT RNACOMO. espirina
Spirin Oleksandr Sergiyovich– acadêmico, diretor do Instituto de Biologia da Academia Russa de Ciências, membro do Presidium da Academia Russa de Ciências.
Em maio de 1953, D. Watson e F. Crick descobriram o princípio da organização estrutural (molecular) da fala do gene - ácido desoxirribonucléico (DNA). A estrutura do DNA deu a chave para o mecanismo da criação exata - reduplicação - da fala do gene. Então vinicla é uma nova ciência - biologia molecular. Bulo formulou o chamado dogma central da biologia molecular: proteínas DNA Yu RNA Yu. O sentido é baseado no fato de que a informação genética é escrita no DNA, realizada na procura de proteínas, mas não sem meio termo, mas por trás de um polímero esporídico adicional - ácido ribonucléico (RNA), e o caminho dos ácidos nucleicos para as proteínas não é negociável. Desta forma, DNA é sintetizado em DNA, garantindo o poder de reduplicação, de forma que a criação do material genético deixou em gerações; O RNA é sintetizado no DNA, resultando em reescrita, transcrição, informação genética na forma de cópias numéricas do RNA; As moléculas de RNA são modelos para a síntese de proteínas - a informação genética é traduzida na forma de lanças polipeptídicas. Em casos especiais, o RNA pode ser reescrito na forma de DNA ("transcrição reversa") e também copiado na forma de RNA (replicação), mas as proteínas nunca podem ser um modelo para ácidos nucleicos (relatado por div.).
Doravante, o próprio DNA significa o declínio dos organismos, de modo que o recrutamento de proteínas, que se encontram em gerações, e o signo a elas associado. A biossíntese da proteína é o processo central da matéria viva, e os ácidos nucléicos são seguros por um lado, por um programa que determina todo o conjunto e especificidade das proteínas que são sintetizadas e, por outro lado, pelo mecanismo de execução precisa dos ciclos em gerações de programas. Doravante, a expectativa de vida na forma clitínica moderna leva à comprovação do mecanismo de biossíntese reduzida de proteínas.
BIOSINTEZ BILKIV
O dogma central da biologia molecular postula apenas formas de transferir a informação genética dos ácidos nucléicos para as proteínas e, então, para o poder e aquele signo de um organismo vivo. O desenvolvimento de mecanismos para a implementação desse caminho por uma década, que se seguiu à formulação do dogma central, expandiu as funções ricamente variadas do RNA, mas foi apenas um portador de informações de genes (DNA) para proteínas e serve como um matriz para a síntese de proteínas.
Na fig. A Figura 1 mostra um esquema geral da biossíntese de proteínas na clitina. RNA mensageiro(RNA mensageiro, RNA mensageiro, mRNA), que codifica proteínas, como já foi dito acima, é apenas uma das três principais classes de RNAs celulares. A principal massa їhnyu (cerca de 80%) para se tornar a próxima classe de RNA - RNA ribossômico, estrutura estrutural yakі utvoryuyut e centros funcionais de partículas sintetizadoras de proteínas universais - ribossomos. O próprio RNA ribossômico é viável - tanto estrutural quanto funcionalmente - moldando máquinas moleculares ultramicroscópicas chamadas ribossomos. Os ribossomos recebem informações genéticas como moléculas de mRNA e, sendo programados pelo restante, geram proteínas no padrão exato de acordo com o programa.
Prote, schob sintetizam proteínas, desprovidas de informações e programas são insuficientes - consumindo material, a partir do qual é possível trabalhar. Material Potik para a síntese de proteínas em ide em ribossomos para a ajuda da terceira classe de RNA celular. RNA de transferência(ARN de transferência, ARN de transferência, ARNt). O fedor se liga covalentemente - aceita - aminoácidos, como um material de brotamento para proteínas, e parece que o aminoacil-tRNA é encontrado no ribossomo. Nos ribossomos, o aminoacil-tRNA interage com códons - combinações de trinucleotídeos - mRNA, após o que ocorre a decodificação dos códons durante o processo de tradução.
ácidos ribonucleicos
Além disso, é possível recrutar o principal RNA celular, o principal processo principal da matéria viva moderna - a biossíntese de proteínas. Ce mRNA, RNA ribossômico e tRNA. O RNA é sintetizado no DNA com a ajuda de enzimas - polimerases de RNA, que induzem a transcrição - reescrevendo as divisões lineares (braços lineares) do DNA de fita dupla na forma de RNA de fita simples. As células do DNA, que codificam proteínas celulares, são reescritas na forma de mRNA, bem como para a síntese de cópias numéricas de RNA ribossômico e tRNA - células especiais do genoma da clitina, para as quais há reescrita intensiva sem tradução adicional de a proteína.
Estrutura química do RNA.
Quimicamente, o RNA já é semelhante ao DNA. Discurso ofensivo - polímeros lineares inteiros de nucleotídeos. O monômero da pele - um nucleotídeo - é uma fosforilação do N-glicosídeo, levando a um excesso de um anel de cinco carbonos - pentose, que carrega um grupo fosfato no grupo hidroxila do quinto átomo de carbono (ligação dobrável) e uma base nitrogenada na o primeiro átomo de carbono (N-glicosina) A diferença química principal entre DNA e RNA naquele que tem um excesso do monômero de RNA - ceribose, e o monômero de DNA - desoxirribose, que é semelhante à ribose, possui um grupo hidroxila diário com outro átomo de carbono (Fig. 2).
 Mal. 2. Fórmulas químicas de ações um dos ribonucleotídeos - uridil ácido (U) e youmu homólogo desoxirribonucleotídeo - ácido timidílico (dT) |
Bases nitrogenadas i no DNA, i no RNA chotiri vide: duas bases purinas - adenina (A) e guanina (G) - duas bases pirimídicas - citosina (C) e uracilo (U) ou timina metilada yogo (T). O uracil é característico dos monômeros de RNA e a timina é característica dos monômeros de DNA, e o RNA e o DNA são igualmente importantes. Os monômeros - ribonucleotídeos de RNA e desoxirribonucleotídeos de DNA - formam uma lança polimérica para a formação adicional de sítios fosfodiéster entre os excedentes sanguíneos (entre o quinto e terceiro átomos de carbono da pentose). Assim, a lança polimérica do ácido nucléico - DNA ou RNA - pode ser representada como um esqueleto linear açúcar-fosfato com bases nitrogenadas como grupo biológico. Estrutura macromolecular do RNA. A diferença macroestrutural fundamental entre os dois tipos de ácidos nucléicos se deve ao fato de o DNA ser uma única hélice, ou seja, uma macromolécula com duas ligações de filamentos poliméricos complementares, espiralados em torno do eixo central (div. [ , ]), e o RNA é um polímero de cadeia simples. Ao mesmo tempo, as interações mútuas dos grupos biológicos - bases nitrogenadas - um a um, e também com fosfatos e hidróxidos do esqueleto açúcar-fosfato são levadas ao ponto em que o polímero de RNA de fita simples se enrola sobre si mesmo e se torce em uma estrutura compacta, semelhante ao dobramento de uma lanceta polipeptídica de uma proteína em um glóbulo compacto. Desta forma, as sequências únicas de nucleotídeos do RNA podem formar extensões únicas de estruturas. |
Anteriormente, a estrutura espacial específica do RNA foi demonstrada ao decifrar a estrutura atômica de um tRNA em 1974. [ , ] (Fig. 3). A formação de uma lanceta polimérica de tRNA, que consiste em monômeros de 76 nucleotídeos, leva à formação de um núcleo globular compacto arqueado, do qual duas cristas são apagadas com um corte reto. Os fedores são espirais de subfios curtos no cluster de DNA, mas também são organizados com a ajuda do suporte mútuo de um e o mesmo tієї e RNA de lanceta. Um deles atua como aceptor de aminoácidos e participa da síntese da proteína lanceta polipeptídica no ribossomo, e o outro é utilizado para interação complementar com o tripleto do mRNA (códon) que codifica no mesmo ribossomo. Somente essa estrutura é especificamente projetada para interagir com a proteína-enzima, que pendura o aminoácido no tRNA, e o ribossomo no processo de tradução, de modo que seja especificamente "reconhecido" por eles.

Mal. 3. Modelos atômicos (canhotos) e esqueléticos (destros) de levedura de tRNA de fenilalanina
A produção de RNAs ribossômicos isolados deu uma aplicação ofensiva e expansiva para a formação de estruturas específicas compactas de polímeros lineares ainda maiores do mesmo tipo. O ribossomo é composto de duas partes nervosas - grandes e pequenas subpartes ribossômicas (subunidades). A subpartícula da pele foi inspirada por um alto RNA polimérico e toda uma série de várias proteínas ribossômicas. O comprimento das lanças no RNA ribossômico é significativo: por exemplo, o RNA da subunidade pequena do ribossomo bacteriano tem mais de 1.500 nucleotídeos e o RNA da subunidade grande tem cerca de 3.000 nucleotídeos. Em sávios, incluindo humanos, o ciRNA é ainda maior - perto de 1900 nucleotídeos e mais de 5000 nucleotídeos em pequenas e grandes subpartes aparentemente.
Foi demonstrado que o RNA ribossômico, água-prata, é isolado de suas proteínas parceiras e retirado de aparência limpa, as próprias estruturas dobram-se espontaneamente em uma estrutura compacta, semelhante em tamanho e forma às subunidades ribossomais]. A forma das subpartículas grandes e pequenas é diferente, e a forma dos RNAs ribossômicos grandes e pequenos é semelhante (Fig. 4). Dessa forma, as lanças lineares do RNA ribossômico se auto-organizam em uma extensão específica de estrutura, o que significa expansão, forma de i, possivelmente, fixação interna de subpartes ribossomais e também de todos os ribossomos.
ARN menor. No mundo, a produção de componentes na clitina viva e nas outras três frações do RNA total da clitina foi estabelecida, mas três dos principais tipos de RNA à direita não estão misturados. Descobriu-se que na natureza existem outros tipos impessoais de RNA. É por isso que o chamamos de "pequeno RNA", que pode abranger até 300 nucleotídeos, muitas vezes com funções desconhecidas. Via de regra, os fedores estão associados a uma ou mais proteínas dekilcom e são apresentados no termo ribonucleoproteínas - "pequeno RNP".
Pequenos RNAs estão presentes em todas as células do clitin, incluindo o citoplasma, núcleo, núcleo, mitocôndrias. A maioria desses pequenos RNPs, que funcionam de alguma forma, participam dos mecanismos de processamento pós-transcricional dos principais tipos de RNA (processamento de RNA) - a transformação de precursores de mRNA em mRNA maduro (splicing), redagulação de mRNA, biogênese de tRNA , maturação do RNA ribossômico. Uma das representações mais abundantes de pequenas espécies de RNP (SRP) na clitina desempenha um papel fundamental no transporte de proteínas que são sintetizadas através da membrana da clitina. Vіdomi vidi pequeno ARN, funções reguladoras scho vykonuyut na tradução. Um pequeno RNA especial entra no depósito da enzima mais importante, conhecida por promover a reduplicação do DNA nas gerações de clitina - a telomerase. Desnecessário dizer que suas dimensões moleculares podem ser comparadas com as dimensões das proteínas globulares celulares. Desta forma, passo a passo, fica claro que o funcionamento das células vivas não é apenas atribuído a diferentes proteínas, mas que são sintetizadas nela, mas também na presença de um rico conjunto de diferentes RNAs, para os quais pequenos RNAs são conhecidos por serem compactos.
Ribozima. Toda vida ativa é motivada pela troca fala-metabolismo, e todas as reações bioquímicas do metabolismo dependem dos recursos necessários para a segurança da vida apenas de catalisadores específicos altamente eficazes, a criação da evolução. Atualizando a riqueza de uma década de bioquímica provou-se que a catálise biológica é constantemente e em toda parte chamada de proteínas, como são chamadas enzimas, ou enzimas. Eixo y 1982-1983 pp. Foi demonstrado que na natureza є espécies de RNA, que, de forma semelhante às proteínas, podem ter atividade catalítica altamente específica [ , ]. Tais catalisadores de RNA foram nomeados ribozimas. A declaração sobre a vinyatkovistity de proteínas na catálise de reações bioquímicas chegou ao fim.
Atualmente, o ribossomo também é comumente visto como uma ribozima. De fato, todos os dados experimentais recentes indicam que a síntese da proteína polipeptídica lanceolada no ribossomo é catalisada pelo RNA ribossômico, e não por proteínas ribossômicas. Foi identificada uma célula catalítica do grande RNA ribossomal, responsável por catalisar a reação de transpeptidação, o que ajuda a aumentar a proteína lanceta polipeptídica no processo de tradução.
Quanto à replicação do DNA viral, esse mecanismo é pouco explorado na reduplicação do material genético - DNA - da própria célula. No caso dos RNAs virais, realizam-se processos que são suprimidos ou às vezes diariamente em células normais, onde todo o RNA é sintetizado apenas no DNA, como em uma matriz. Quando infectado com vírus de RNA, a situação pode ser dupla. Em alguns casos, no RNA viral, como em um molde, o DNA é sintetizado ("transcrição reversa"), e cópias numéricas do RNA viral são transcritas neste DNA. Em outro, o mais importante para nós ataques ao RNA viral, é sintetizada uma lanceta de RNA complementar, que serve de molde para a síntese - replicação - de novas cópias do RNA viral. Desta forma, durante a infecção com vírus de RNA, o princípio da formação do RNA é realizado para determinar a criação de sua própria estrutura de água, como é o caso do DNA.
Multifuncionalidade do RNA. Dado o conhecimento sobre as funções do RNA, é possível falar sobre a funcionalidade não primária deste polímero na natureza viva. É possível dar um turno ofensivo das funções principais do ARN.
Função replicativa genética: capacidade estrutural de copiar (replicar) sequências lineares de nucleotídeos por meio de sequências complementares. A função é realizada em infecções virais e é semelhante à principal função do DNA na vida dos organismos clitínicos - a reduplicação do material genético.Como um todo, o RNA está diante de nós com um polímero maravilhoso, o que, teria sido, não para o tempo de evolução do Mundo-Todo, nem para o intelecto do Criador, não bastaria falar sobre vinho de ioga. Como é possível para bulo bachiti, o RNA é capaz de violar as funções da vida fundamentalmente importante dos polímeros - DNA e proteínas. Não é de admirar que a ciência tenha recebido alimento: por que essa base autossuficiente do mundo do RNA não poderia ser destruída pelo surgimento da vida em sua forma moderna de DNA-proteína?Função de codificação: programação da síntese de proteínas por sequências lineares de nucleótidos. Esta é a mesma função que o DNA tem. I no DNA, i no RNA, um mesmo trio de nucleotídeos codifica 20 aminoácidos da proteína, e a sequência do trio nas lanças do ácido nucléico é um programa para a ordenação sequencial de 20 tipos de aminoácidos em a lança polipeptídica da proteína.
Função de criação de estrutura: formando estruturas triviais únicas. Pequenas moléculas de RNA compactamente dobradas são basicamente semelhantes às estruturas trivímeras de proteínas globulares, e moléculas de RNA maiores podem formar partículas biológicas ou núcleos maiores.
A função de reconhecimento: interações espaciais altamente específicas com outras macromoléculas (incluindo proteínas e outros RNAs) e pequenos ligantes. Essa função, talvez, seja a principal das proteínas. Baseia-se na classificação única zgortatisya do polímero zdatnostі e na moldagem de estruturas trivimіrnі específicas. A função de reconhecimento é a base da catálise específica.
Função catalítica: catálise específica de reações químicas por ribozimas. Esta função é semelhante à função enzimática das proteínas-enzimas.
VIVENDO A VIDA
Teoria da proteína-coacervato de Oparin. Talvez, ciência pershu, uma teoria bem pensada da jornada da vida por um caminho abiogênico tenha sido sugerida pelo bioquímico A.I. Oparin na década de 20 do século passado [ , ]. A teoria baseava-se no fato de que tudo partiu das proteínas e na possibilidade de síntese química espontânea de monômeros de proteínas - aminoácidos - e polímeros semelhantes a proteínas (polipeptídeos) de forma abiogênica. A publicação da teoria estimulou experimentos numéricos em vários laboratórios do mundo, pois mostraram a realidade de tal síntese em mentes de peças. A teoria de Shvidko tornou-se amplamente aceita e extremamente popular.
O principal postulado її foi aqueles que espontaneamente culpados no "caldo" primário das lajes semelhantes a proteínas foram combinados "em gotas coacervadas - cremação de água da coluna do sistema (cinzas), dovkilla, їi compartimentos assim, então yak dekі bilkovo subluxis spluoks Koarvasnaya Krapel poderia ter um séssil de ativo ativo, então a passagem da peste da porcaria sincera é um róseo -ninnica do asyelasi, e o zeroinel do pronome é Coacervatus, que assimila, cresce e se multiplica sob a pele, parecendo um protótipo de célula viva (Fig. 5).
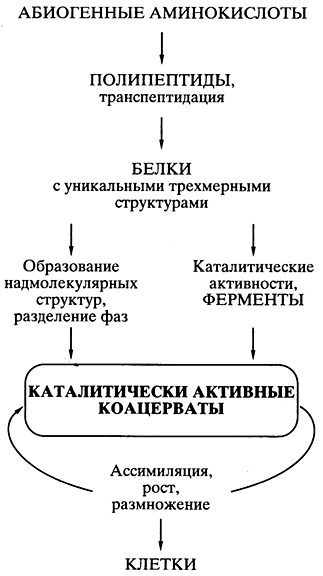
Mal. 5. Representação esquemática dos caminhos da jornada da vida
de acordo com a teoria da proteína-coacervato de A.I. Oparina
Tudo foi bem pensado e cientificamente fundamentado na teoria, exceto por um problema, por muito tempo os olhos de todos os fahivts da galeria ficaram achatados por muito tempo. Якщо спонтанно, шляхом випадкових безматричних синтезів у коацерваті виникали поодинокі вдалі конструкції білкових молекул (наприклад, ефективні каталізатори, що забезпечують перевагу даному коацервату в зростанні та розмноженні), то як вони могли копіюватися для поширення всередині коацервату, а тим більше для передачі коацерватам- Теорія apareceu intempestivamente para propagar o problema da criação exata - no meio do coacervado e nas gerações - de estruturas únicas de proteínas efetivas, que surgiram do nada.
RNA leve como uma frente vida cotidiana. Conhecimento acumulado sobre Código genético, ácidos nucléicos e a biossíntese de proteínas levaram à confirmação de uma ideia fundamentalmente nova sobre o TOM, de que tudo começou não a partir de proteínas, mas de RNA [-]. Нуклеїнові кислоти є єдиним типом біологічних полімерів, макромолекулярна структура яких завдяки принципу комплементарності при синтезі нових ланцюгів (докладніше див.) забезпечує можливість копіювання власної лінійної послідовності мономерних ланок, іншими словами, можливість відтворення (реплікації) полімеру, його мікроструктури. Portanto, apenas os ácidos nucléicos, mas não as proteínas, podem ser um material genético, ou seja, moléculas que repetem sua microestrutura específica em gerações.
É o baixo nível do próprio RNA, e não o DNA, que pode ser o primeiro material genético.
Primeiro, i na síntese química, i nas reações bioquímicas, os ribonucleotídeos são convertidos em desoxirribonucleotídeos; Os desoxirribonucleotídeos são produtos da modificação dos ribonucleotídeos (div. Fig. 2).
De uma maneira diferente nos processos universais mais recentes do metabolismo da vida, os ribonucleotídeos, em vez dos desoxirribonucleotídeos, estão amplamente representados, incluindo as principais fontes de energia do tipo ribonucleosídeo-polifosfato (ATP pobre).
Em terceiro lugar, A replicação do RNA pode ser realizada sem qualquer envolvimento do DNA, e o mecanismo de reduplicação do DNA pode ser encontrado no mundo vivo para influenciar o papel geral do iniciador de RNA na iniciação da síntese da lança do DNA.
Em quarto lugar possuindo toda a mesma matriz e funções genéticas, como DNA, RNA, também são capazes de funções baixas, proteínas poderosas, incluindo catálise de reações químicas. Dessa forma, é possível considerar o DNA como uma nova abordagem evolutiva - como uma modificação do RNA, especializado para fins de funções vitoriosas e criação de cópias únicas de genes no depósito do genoma do clitin sem a participação ininterrupta da biossíntese .
Além disso, assim que o RNA cataliticamente ativo foi usado, a ideia da primazia do RNA na vida deu um forte impulso ao desenvolvimento e o conceito foi formulado mundo autossuficiente de RNA, scho soprando a vida atual [ , ]. Um esquema possível do ARN viknennia mostra-se no figo. 6.
A síntese abiogênica de ribonucleotídeos na mesma associação covalente no oligômero e polímero do tipo RNA pôde ser observada aproximadamente nas mesmas mentes e nas mesmas condições químicas que foram postuladas para a síntese de aminoácidos e polipeptídeos. Recentemente O.B. Chetverin e seus colegas (Instituto de Proteína RAS) mostraram experimentalmente que os polirribonucleotídeos (RNA) são utilizados no ambiente aquático primário antes da recombinação espontânea, de modo que a troca de lanceolados pela via da transesterificação. A troca de nervuras lancetas curtas por longo tempo é responsável pela redução dos polirribonucleotídeos (RNA), e a própria recombinação é semelhante à diversidade estrutural dessas moléculas. Entre eles, as moléculas de RNA cataliticamente ativas podem ser culpadas.
Provocar o aparecimento de moléculas únicas de RNA, que poderiam catalisar a polimerização de ribonucleotídeos ou o splicing de oligonucleotídeos na lança complementar como na matriz [ ], significava a formação do mecanismo de replicação do RNA. A replicação dos próprios catalisadores de RNA (ribozimas) tem pouca importância em causar populações de RNA autorreplicantes. Produzindo suas próprias cópias de RNA se multiplicaram. Perdões inevitáveis na cópia (mutação) e recombinação em populações de RNA, que se autorreplicam, criaram cada vez mais diversidade em todo o mundo. Nesta classificação, a transferência do antigo mundo do RNA - ce "um mundo biológico autossuficiente, no qual as moléculas de RNA funcionam como material genético e como catalisadores semelhantes a enzimas" .
Vindicação para a biossíntese de proteínas. Далі на основі світу РНК мало відбуватися становлення механізмів біосинтезу білка, поява різноманітних білків із успадкованою структурою та властивостями, компартменталізація систем біосинтезу білка та білкових наборів, можливо, у формі коацерватів та еволюція останніх у клітинні структури – живі клітини (див. рис. 6) . ).
O problema da transição do antigo mundo do RNA para o atual mundo da síntese de proteínas é importante para uma solução fundamentalmente teórica. A possibilidade de síntese abiogênica de polipeptídeos e falas protéicas não ajuda a resolver o problema, não há indícios de nenhum caminho específico, como a síntese de revestimentos de bibuty mig a partir de RNA e sob controle genético. O controle genético da síntese de polipeptídeos e proteínas se desenvolve independentemente da síntese abiogênica primária, à sua maneira, com o aprimoramento do RNA já existente. Na literatura, algumas hipóteses foram propostas para o mecanismo moderno de biossíntese de proteínas no mundo do RNA, mas, talvez, possam ser vistas como pensadas em detalhes e sem um olhar de capacidades físicas e químicas. Apresento minha versão do processo de evolução e especialização do RNA, que levou ao fim do aparato de biossíntese de proteínas (pequeno 7), mas não pretendo terminar.
Foi proposto um esquema hipotético para vingar duzentos momentos, que parecem importantes.
Primeiro, Postula-se que os oligorribonucleotídeos sintetizados abiogenicamente foram ativamente recombinados por trás do mecanismo adicional de transesterificação não enzimática espontânea, levando à redução de lancetas de RNA e dando origem à sua diversidade. Da mesma forma, na população de oligonucleotídeos e polinucleotídeos, tanto espécies de RNA cataliticamente ativas (ribozimas) como outras espécies de RNA com funções especializadas podem aparecer (Fig. 7). Além disso, a recombinação não enzimática de oligonucleotídeos que se ligam complementarmente ao molde polinucleotídico poderia assegurar a fusão (splicing) de fragmentos complementares a este molde em uma única lança. Desta forma, em vez de catalisar a polimerização de mononucleótidos, pode ser efectuada a cópia primária (multiplicação) do ARN. Assumindo que eram ribozimas que tinham pouca atividade de polimerase, então a eficiência (precisão, flexibilidade e produtividade) de copiar em complementares. matrizes são pequenas e crescem significativamente.

Mal. 7. Esquema de evolução e especialização de moléculas de RNA
no processo de transição do antigo mundo do RNA para para o mundo atual
Biossíntese de proteínas geneticamente determinadas
Outro Um ponto importante em minha versão é que o primeiro aparato para a biossíntese de proteínas apareceu na erradicação de muitas espécies de RNA especializado antes do surgimento do aparato para replicação enzimática (polimerase) de material genético - RNA e DNA. Seu aparato primário, incluindo o RNA proribossomal cataliticamente ativo, tem baixa atividade de peptidiltransferase; recrutamento de pró-tRNAs que se ligam especificamente a aminoácidos ou peptídeos curtos; outro RNA proribossomal, interagindo simultaneamente com RNA proribossomal catalítico, pro-mRNA e pro-tRNA (div. Fig. 7). Tal sistema já poderia sintetizar polipeptídeos lanceuge através de uma reação de transpeptidação catalisada. Entre as outras proteínas cataliticamente ativas - as enzimas primárias (enzimas) - também havia proteínas que catalisam a polimerização de nucleotídeos - réplicas ou polimerases NK.
Vtim, talvez, qual é a hipótese sobre luz antiga O RNA, como líder do mundo vivo de hoje, não pode ser suficiente para fundamentar o problema principal - uma descrição cientificamente plausível do mecanismo de transição do RNA e replicação para a biossíntese de proteínas. A hipótese alternativa de A.D. Альтштейна (Інститут біології гена РАН), в якій постулюється, що реплікація генетичного матеріалу та його трансляція - синтез білка - виникали і еволюціонували одночасно і поєднано, починаючи з взаємодії абіогенно синтезованих олігонуклеотидів і аміноацил-нуклеотидилатів - змішаних ангідридів. Infelizmente, o cossaco já está chegando ... ( "I Shahrazad pegou a ferida e ela fixou a permissão para promo".)
Literatura
1. Watson JD, Crick F.H.C. Estrutura molecular dos ácidos nucleicos // Natureza. 1953. V. 171. P. 738-740. 2. Watson JD, Crick F.H.C. Implicações genéticas da estrutura dos ácidos nucléicos desoxirribose // Nature 1953 V. 171. P. 964-967. 3. Spirina A.S. Biologia moderna e segurança biológica // Boletim da Academia Russa de Ciências. 1997. Nº 7. 4. Spirina A.S. Sobre a estrutura macromolecular da química ribonuclear natural de alto polímero // Journal of Molecular Biology. 1960. V. 2. P. 436-446. 5. Kirn S.H., Suddath F.L., Quigley GJ. isso em. Estrutura terciária tridimensional do RNA de transferência de fenilalanina de levedura // Science. 1974. V. 185. P. 435-40. 6. Robertas J.D., Ladner J.E., Finch J.T. isso em. Estrutura do tRNA de fenilalanina de levedura em resolução de 3 A // Nature. 1974. V. 250. P. 546-551. 7. Vasiliev V.D., Serdyuk I.N., Gudkov A.T., SPIRin A.S. Auto-organização do RNA ribossômico // Estrutura, Função e Genética dos Ribossomos / Eds. Hardesty B. e Kramer G. Nova York: Springer-Verlag, 1986, pp. 129-142. 8. Baserga SJ., Steitz J.A. Raznomanіtny svіt malikh ribo-nucleoproteínas // The RNA World / Eds. Gesteland R.F. e Atkins J. F. Nova York: Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1993, pp. 359-381. 9. Kruger K., Grabowski PJ., Zaug AJ. isso em. ARN de auto-splicing: Autoexcisão e autociclização da sequência interveniente do ARN ribossómico de Tetrahymena// célula. 1982. V. 31. P. 147-157. 10. Guerrier-Takada C., Gardiner K., Marsh T. et al. A fração de RNA ribonucleases P é uma subunidade catalítica da enzima // Cell. 1983. V. 35. P. 849-857. 11. Oparin A.I. Caminhando a vida. M: Moscou Robotnik, 1924. 12. Oparin A.I. Vinnyknennya vida da Terra (3º tipo). M: Vista da Academia de Ciências da SRSR, 1957. 13. Woese S. Evolução do código genético // O Código Genético. Nova York: Harper & Row, 1967, pp. 179-195. 14. Crick F.H.C. origem do código genético // Journal of Molecular Biology. 1968. V. 38. P. 367-379. 15. Orgel L.E. Evolução do aparelho genético // Journal of Molecular Biology. 1968. V. 38. P. 381-393. 16. Gilberto W. O mundo do RNA // Natureza. 1986. V 319 P. 618. 17. Joyce GF, Orgel L.E. Perspectivas para a compreensão da origem do mundo do RNA // The RNA World / Eds. Gesteland R.F. e Atkins J. F. Nova York: Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1993 P 1-25. 18. Chetverina H.V., Demidenko A.A., Ugarov V.I., Chetverin A.B. Reorganização espontânea em sequências de RNA // FEBS Letters. 1999. V. 450. P. 89-94. 19. Bartel D.P., Szostak J.W. Isolamento de novas ribozimas a partir de um grande conjunto de sequências aleatórias // Science. 1993. V. 261. P. 1411-1418. 20. Ekland E.H., Bartel D.P. Polimerização de RNA catalisada por RNA com trifosfatos de nucleosídeos // Nature. 1996 V. 382. P. 373-376. 21. Orgel L.E. origem da vida - Revisão de fatos e especulações // Tendências em Ciências Bioquímicas. 1998. V. 23. pág. 491-495. 22. Altstein A. D. Sistema genético Pohodzhennya: a hipótese da descendência // Biologia Molecular. 1987. T. 21. S. 309-322.








