در تمام سلول های زنده، پروتئین ها توسط ریبوزوم ها سنتز می شوند . ریبوزوم یک ماکرومولکول بزرگ با ساختار یک چهارم نامتقارن چین خورده است که از اسیدهای ریبونوکلئیک (RNA ریبوزومی) و پروتئین ها ساخته شده است. به منظور سنتز پروتئین ها، ریبوزوم با:
1. برنامه ای که ترتیب پردازش اسیدهای آمینه اضافی پروتئین پلی پپتیدی را تعیین می کند.
2. مواد اسید آمینه، که از آن پروتئین وجود خواهد داشت.
3. انرژی.
ریبوزوم خود یک عملکرد کاتالیزوری (آنزیمی) دارد که مسئول ایجاد پیوندهای پپتیدی و احتمالاً پلیمریزاسیون باقی مانده اسیدهای آمینه در پروتئین های پلی پپتیدی است.
برنامه ای که ترتیب اسیدهای آمینه اضافی را در لانکوس پلی پپتیدی پروتئین تنظیم می کند به صورت دی اکسی ریبونوکلئیک اسید (DNA) یعنی ژنوم سلول ظاهر می شود. رشته های RNA سنتز شده مکمل یکی از رشته های DNA هستند و به این ترتیب توالی دئوکسی ریبونوکلئوتید رشته DNA دیگر را در توالی ریبونوکلئوتیدی خود به طور دقیق بازتولید می کنند. فرآیند کپی کردن یک ژن که توسط آنزیم RNA پلیمراز پردازش می شود، رونویسی نامیده می شود. در طول سنتز RNA، به ویژه در سلولهای یوکاریوتی، میتواند تحت تعدادی تغییرات اضافی به نام پردازش قرار گیرد که در طی آن میتواند محتوای توالی نوکلئوتیدی را تغییر دهد. RNA آزاد شده به سمت ریبوزوم های هسته برنامه می رود، که توالی اسید آمینه را در پروتئین سنتز شده تعیین می کند. به آن RNA اطلاعات یا «پیامرسان» (mRNA) میگویند. بنابراین، خود رونویسی ژن ها و ایجاد mRNA جریان اطلاعات از DNA به ریبوزوم ها را تضمین می کند.
ماده بلال که پروتئین تولید می کند، حاوی اسیدهای آمینه است. با این حال، اسیدهای آمینه آزاد توسط ریبوزوم جذب نمی شوند. برای اینکه به عنوان یک سوبسترا برای ریبوزوم عمل کند، اسید آمینه باید از طریق شکاف حاصل از ATP فعال شود و توسط یک مولکول RNA خاص به نام انتقال یا انتقال پذیرفته شود (به صورت کووالانسی پذیرفته شود). ї RNA (tRNA)، که به آنزیم سنتز کمک می کند. aminoacyl-tRNA به عنوان بستری برای سنتز پروتئین از ریبوزوم حذف می شود. علاوه بر این، انرژی پیوند شیمیایی بین باقی مانده اسید آمینه و tRNA برای واکنش پیوند پپتیدی در ریبوزوم استفاده می شود. بنابراین، فعال شدن اسیدهای آمینه و ایجاد aminoacyl-tRNA جریان مواد و انرژی برای سنتز پروتئین ریبوزومی را تضمین می کند.
این سه جریان (اطلاعات، مواد و انرژی) در ریبوزوم همگرا می شوند. ریبوزوم با دریافت آنها، ترجمه یا ترجمه اطلاعات ژنتیکی را از توالی نوکلئوتیدی mRNA به توالی اسید آمینه پروتئین پلی پپتیدی سنتز شده انجام می دهد. همانطور که این به صورت مولکولی بیان می شود، ریبوزوم به طور متوالی DNA mRNA را اسکن می کند (بعد از آن فرو می ریزد) و سپس به طور متوالی aminoacyl-tRNA را از وسط انتخاب می کند، که منجر به اختصاصی بودن باقیمانده آمینواسیل در انتخاب توسط aminoacyl-tRNA می شود. ریبوزوم به شدت توسط ویژگی ترکیب تعیین می شود. قطعه mRNA بنابراین، مشکل کد ژنتیکی پیش میآید: چه ترکیبی از نوکلئوتیدها تعیین میشود، یعنی چه چیزی پوست را با 20 اسید آمینه کد میکند، که مولکولهای پروتئین از آن تشکیل میشوند؟
جریان ریبوزوم از طریق ریبوزوم mRNA (یا به عبارت دیگر، عبور mRNA لانجوگ از ریبوزوم) یک ترتیب ساعتی دقیقی را برای ورود به ریبوزوم آمینواسیل-tRNA های مختلف از زیرگونه به ترتیب تعیین می کند. چرخش ترکیبات نوکلئوتیدی که mRNA سالم را کد می کند. بقایای آمینواسیل آمینواسیل-tRNA مونتاژ شده بلافاصله به صورت کووالانسی توسط ریبوزوم به نیزه پلی پپتیدی در حال رشد متصل می شود. tRNA دی اسیله شده به ریبوزوم های گونه های مختلف متصل می شود. بنابراین، به طور متوالی، گام به گام، لنست پلی پپتیدی پروتئین تشکیل می شود (شکل 1).
بیوسنتز پروتئین.
متابولیسم پلاستیک (همسان سازی و آنابولیسم) مجموعه ای از واکنش های سنتز بیولوژیکی است. این نوع مبادله با ذات خود خوانده می شود: از سخنرانی ها، مانند سخنرانی های مشتری، سخنرانی هایی شبیه به سخنرانی های مشتری ایجاد می شود.
بیایید به یکی از مهمترین اشکال متابولیسم پلاستیک - بیوسنتز پروتئین نگاه کنیم. بیوسنتز پروتئین هادر تمام سلول های مربوط به یوکاریوت ها وجود دارد. اطلاعات مربوط به ساختار اولیه (ترتیب اسید آمینه) یک مولکول پروتئین توسط توالی نوکلئوتیدها در پسوند مولکول DNA - ژن ها رمزگذاری می شود.
این ژن بخشی از مولکول DNA است که به معنای ترتیب اسیدهای آمینه در مولکول پروتئین است. همچنین نوکلئوتیدهای موجود در ژن ترتیب اسیدهای آمینه را در پلی پپتید تعیین می کنند. این ساختار اولیه ای است که سایر ساختارها، قدرت ها و عملکردهای مولکول پروتئین در آن قرار دارند.
سیستم ثبت اطلاعات ژنتیکی در DNA (و RNA) با توجه به توالی دقیق نوکلئوتیدها، کد ژنتیکی نامیده می شود. توبتو. یک واحد کد ژنتیکی (کدون) سه گانه نوکلئوتید در DNA یا RNA است که یک اسید آمینه را کد می کند.
کل کد ژنتیکی شامل 64 کدون است که 61 کد کد و 3 کد غیر کد کننده هستند (کدون های پایان دهنده که نشان دهنده تکمیل فرآیند ترجمه هستند).
کدون پایان دهنده i - RNA: UAA، UAG، UGA، در DNA: ATT، ATC، ACC.
آغاز فرآیند ترجمه کدون آغازگر (AUG، DNA - TAC) است که آمینو اسید متیونین را کد می کند. این کدون اولین کدونی است که وارد ریبوزوم می شود. سپس متیونین از آنجایی که به عنوان اولین اسید آمینه این پروتئین منتقل نمی شود، جدا می شود.
کد ژنتیکی مشخصه قدرت است.
1. جهانی بودن - کد برای همه موجودات یکسان است. همان سه گانه (کدون) در هر ارگانیسمی همان اسید آمینه را رمزگذاری می کند.
2. ویژگی - هر کدون فقط یک اسید آمینه را کد می کند.
3. Genericity - اکثر اسیدهای آمینه را می توان توسط چندین کدون رمزگذاری کرد. دو اسید آمینه مقصر هستند - متیونین و تریپتوفان که تنها با یک نوع کدون مرتبط هستند.
4. بین ژن ها "علائم تقسیم" وجود دارد - سه سه قلو خاص (UAA، UAG، UGA)، که هر کدام نشان دهنده سنتز لانژوگ پلی پپتیدی است.
5. هیچ "علائم تقسیمی" در وسط ژن وجود ندارد.
برای اینکه یک پروتئین سنتز شود، اطلاعات مربوط به توالی نوکلئوتیدها در ساختار اولیه آن ممکن است به ریبوزوم ها تحویل داده شود. این فرآیند شامل دو مرحله است - رونویسی و ترجمه.
رونویسی(بازنویسی) اطلاعات از طریق سنتز یک طرف مولکول DNA یک مولکول RNA یک طرفه به دست می آید که توالی نوکلئوتیدهای آن دقیقاً با دنباله نوکلئوتیدهای ماتریکس - مولکول DNA پلی نوکلئوتیدی مطابقت دارد.
Vaughn (i - RNA) واسطه ای است که اطلاعات را از DNA به محل تجمع مولکول های پروتئین در ریبوزوم منتقل می کند. سنتز RNA (رونویسی) در مراحل اولیه انتظار می رود. یک آنزیم (RNA پلیمراز) رشته DNA را شکافته و یکی از رشته ها (کد کننده) نوکلئوتیدهای RNA را بر اساس اصل مکملیت تولید می کند. مولکول RNA به این روش سنتز می شود (سنتز الگو) و وارد سیتوپلاسم می شود و زیر واحدهای کوچکی از ریبوزوم ها در یک انتها رشته می شوند.
مرحله دیگر در بیوسنتز پروتئین است پخش- این ترجمه توالی نوکلئوتیدها در یک مولکول است و - RNA دنباله اسیدهای آمینه در یک پلی پپتید است. در پروکاریوتها که هسته تشکیلشده ندارند، ریبوزومها میتوانند پس از جدا شدن از DNA به مولکول تازه ایجاد شده و آنزیم RNA متصل شوند یا منجر به تکمیل کامل سنتز شوند. در یوکاریوت ها، i-RNA از طریق پوشش هسته ای به سیتوپلاسم منتقل می شود. انتقال توسط پروتئین های خاصی انجام می شود که با مولکول و RNA کمپلکس ایجاد می کنند. آنها علاوه بر عملکرد انتقال این پروتئین ها، i-RNA را از عمل می گیرند که باعث از بین رفتن آنزیم های سیتوپلاسمی می شود.
در سیتوپلاسم، در یک انتهای RNA (و در همان انتهایی که سنتز مولکول در هسته شروع می شود)، یک ریبوزوم وارد می شود و سنتز پلی پپتید آغاز می شود. هنگامی که یک ریبوزوم با یک مولکول RNA بارگذاری می شود، سه گانه پس از سه گانه ترجمه می شود و متوالی اسیدهای آمینه را به انتهای بالغ حلقه پلی پپتیدی اضافه می کند. هویت دقیق کد اسید آمینه به سه گانه و RNA توسط tRNA تضمین می شود.
انتقال RNA (tRNA) اسیدهای آمینه را به زیر واحد بزرگ ریبوزوم می آورد. مولکول t-RNA دارای یک پیکربندی تا شده است. در این کرت ها، پیوندهای آبی بین نوکلئوتیدهای مکمل تشکیل می شود و شکل مولکول شبیه یک برگ پایدار است. در بالای انبساط یک سه گانه از نوکلئوتیدهای قوی (آنتیکودون) وجود دارد که نشان دهنده اسید آمینه اصلی است و پایه جایی است که این اسید آمینه متصل شده است (شکل 1).
کم اهمیت 1. طرح RNA انتقال آینده: 1 - پیوندهای آبی. 2 – آنتی کدون رتبه سوم اتصال اسید آمینه
T-RNA پوست می تواند اسید آمینه خود را منتقل کند. T-RNA توسط آنزیم های خاصی فعال می شود، اسید آمینه آن را می پذیرد و به ریبوزوم منتقل می کند. در وسط ریبوزوم در حال حاضر تنها دو کدون i-RNA وجود دارد. از آنجایی که آنتی کدون t-RNA مکمل کدون i-RNA است، t-RNA بلافاصله به اسید آمینه به i-RNA اضافه می شود. قبل از کدون دیگری، t-RNA دیگری اضافه می شود که اسید آمینه خود را حمل می کند. اسیدهای آمینه توسط زیر واحد بزرگ ریبوزوم سنتز می شوند و با کمک آنزیم ها پیوند پپتیدی بین آنها برقرار می شود. در همان زمان، پیوند بین اولین اسید آمینه و t-RNA و t-RNA با ریبوزوم های پشت اولین اسید آمینه تشکیل می شود. ریبوزوم هر بار یک سه قلو را حرکت می دهد و این روند تکرار می شود. به این ترتیب، یک مولکول پلی پپتیدی به تدریج رشد می کند، که در آن اسیدهای آمینه در همان نوع با ترتیب سه قلوهایی که آنها را رمزگذاری می کنند (سنتز الگو) تشکیل می شوند (شکل 2).

کم اهمیت 2. طرح بی سنتز پروتئین: 1 - i-RNA; 2 - زیر واحدهای ریبوزومی; 3 - t-RNA با اسیدهای آمینه. 4 - t-RNA بدون اسیدهای آمینه. 5 - پلی پپتید 6 - کدون ta-RNA; tRNA 7 آنتی کودون.
یک ریبوزوم قادر به سنتز یک لانست پلی پپتیدی جدید است. پروت، اغلب یک مولکول i-RNA دسته ای از ریبوزوم ها را فرو می ریزد. به چنین کمپلکس هایی پلی ریبوزوم می گویند. پس از اتمام سنتز، نیزه پلی پپتیدی به یک ماتریکس - یک مولکول i-RNA تبدیل می شود، به شکل یک مارپیچ تا می شود و یک ساختار ثانویه (ثانویه، سوم یا چهارم) را تشکیل می دهد. ریبوزوم ها بسیار کارآمد عمل می کنند: ریبوزوم باکتریایی ظرف یک ثانیه یک لنست پلی پپتیدی حاوی 20 اسید آمینه را هضم می کند.
بیوسنتز پروتئین ها (پلی پپتیدها) فرآیندی بسیار پیچیده و شگفت انگیز است. بیوسنتز پروتئین به طور فعال در تمام اندام ها و بافت ها از جمله گلبول های قرمز اتفاق می افتد. بسیاری از سلولها پروتئینها را برای «صادرات» سنتز میکنند (سلولهای کبد، سابها)، و در این مورد حتی تعداد زیادی ریبوزوم دارند. در کشت حیوانات، تعداد ریبوزوم ها به 105 می رسد و قطر ریبوزوم به 20 نانومتر می رسد.
فرآیند سنتز پروتئین در وسط سلولهای روی سطح ریبوزومها اتفاق میافتد که کمپلکسهایی از دو زیر واحد با ثابت تهنشینی 60S و 40S هستند که بهعنوان یک کل واحد عمل میکنند. پروتئین های ریبوزوم 30-35٪ و RNA ریبوزومی - 65-70٪ می شوند. ریبوزوم ها به دو بخش آمینواسیل و پپتیدیل تقسیم می شوند. اولی برای تثبیت مجموعه اسید آمینه فعال و tRNA که روی ریبوزوم قرار دارد عمل می کند و دیگری لانست پلی پپتیدی متصل به tRNA دیگر را ثابت می کند. زیر واحدهای ریبوزوم در هسته روی یک الگوی DNA سنتز می شوند.
ماهیت فرآیند سنتز پروتئین توسط نمودار نشان داده شده است:
سیستم سنتز پروتئین شامل ریبوزوم ها، اسیدهای نوکلئیک، مجموعه ای از 20 اسید آمینه، آنزیم های مختلف، ATP، GTP، یون های منیزیم و تقریبا 200 فاکتور مختلف پروتئین غیر کاتالیزوری است.
یک مولکول پروتئین گروه بزرگی از اسیدهای آمینه است که به طور متوسط شامل 100 تا 500 اسید آمینه است. برنامه سنتز پروتئین پوست در مولکول دی اکسی ریبونوکلئیک اسید (DNA) ذخیره می شود. مولکول DNA یک پلیمر است که مونومرهای آن نوکلئوتید هستند. توالی بازهای نیتروژنی در یک مولکول DNA، توالی اسیدهای آمینه را در یک مولکول پروتئین تعیین می کند.
مولکول DNA دارای چهار نوع باز نیتروژنی است: آدنین (A)، گوانین (G)، سیتوزین (C) و تیمین (T). دنباله ای از سه جانشین (سه گانه) برای تشکیل یک کدون که یک اسید آمینه را نشان می دهد استفاده می شود.
اسیدهای نوکلئیک - DNA و RNA - اجزای اتصال دهنده بیوسنتز پروتئین هستند. DNA مسئول ذخیره اطلاعات ژنتیکی است، در حالی که RNA مسئول انتقال این اطلاعات و پیاده سازی آن در مولکول های پروتئین است. می توان بیان کرد که وظیفه اصلی DNA حفظ ژنوتیپ و RNA حفظ ژنوتیپ ویروسی است.
در کشت، RNA ریبوزومی (rRNA) اهمیت بیشتری دارد. rRNA را می توان مارپیچی کرد و حاوی نوکلئوتیدهای اصلاح شده (مثلاً 2-متیل ریبوز) باشد. rRNA تقریباً 80٪ از مقدار کل RNA در سلول ها را تشکیل می دهد. نوع دیگری از RNA در سلول ها با RNA انتقالی (tRNA) نشان داده می شود که مانند سایر انواع RNA در هسته سنتز می شود. این قسمت 10-15 درصد از کل مقدار RNA در سلول ها را تشکیل می دهد. بیش از 60 tRNA مختلف شناسایی شده است. همچنین، انتقال اسیدهای آمینه به تعدادی از tRNA های مختلف بستگی دارد. یک tRNA خاص برای اسیدهای آمینه پوست در سلول ها وجود دارد. مولکول های tRNA یکسان هستند. ساختار آنها 75-93 ریبونوکلئید دارد.
اسید آمینه به گروه 3-OH قوی مونونوکلئوتید انتهایی tRNA اضافه می شود که ابتدا با اسید آدنیلیک نشان داده می شود. tRNA یک عنصر مهم دیگر دارد - یک آنتی کدون که علاوه بر آن کمپلکس اسید آمینه و tRNA توالی سه نوکلئوتید را در الگوی RNA (کدون) تعیین می کند. آنتی کدون و کدون به طور مکمل به اتصالات آب متصل هستند.
از آنجایی که ما اطلاعات انفجاری را در سلول حمل می کنیم، DNA که در هسته موجود است، و سنتز پروتئین در سیتوپلاسم انجام می شود، پس یک واسطه آوازخوان وجود دارد که این اطلاعات را به سیتوپلاسم سلول منتقل می کند. Itini. واسطه اطلاعات RNA پیام رسان (mRNA) بود. قبل از mRNA 2٪ از محتوای سلولی سلول های RNA وجود دارد. مولکولهای mRNA یافت شده (شامل حداکثر 5 هزار نوکلئوتید). mRNA همچنین حاوی چندین نوع باز نیتروژنی است. سه مورد از آنها (A، G، C) مانند DNA هستند و چهارمی اوراسیل است.
اطلاعات رمزگذاری شده در mRNA برای سنتز مولکول پروتئینی که روی ریبوزوم ها تولید می شود ضروری است. سنتز mRNA در هسته سلول بسیار سریع است که به بیوسنتز فعال مولکول های پروتئین نیاز دارد. mRNA روی یکی از رشته های DNA هسته ایجاد می شود. در این مورد، ساختار دو رشته ای DNA باز می شود و سنتز DNA RNA پلیمراز از اصل مکمل بودن پیروی می کند:

طرحی برای سنتز mRNA
اصل مکمل بودن به این معنی است که آدنین روی مارپیچ DNA توسط mRNA اوراسیل، تیمین توسط آدنین و گوانین توسط سیتوزین نشان داده می شود. همچنین mRNA اطلاعات را از DNA می خواند.
بنابراین مرحله DNA - RNA به معنای سنتز یک مولکول mRNA است که در آن توالی نوکلئوتیدی مکمل توالی DNA (ژن) است. این فرآیند رونویسی نامیده می شود. سپس mRNA به ریبوزوم می رسد و به زیر واحدهای آن می پیوندد. یک مولکول mRNA به چندین ریبوزوم به طور همزمان ثابت می شود که اصطلاحاً پلی زوم نامیده می شود. وجود پلی زوم ها باعث افزایش کارایی و سیالیت تولید mRNA می شود.
سنتز لانست پلی پپتیدی بر روی یک الگوی mRNA انجام می شود. فرآیند انتقال از mRNA به پروتئین ترجمه نامیده می شود. مرحله "RNA -> پروتئین" فرآیند سنتز پروتئین است که به mRNA هدایت می شود. بنابراین، انتقال اطلاعات همیشه مستقیماً از طریق DNA - RNA - پروتئین انجام می شود.
فرآیند ترجمه شامل مراحل زیر است:
این آنزیم در تثبیت اسید آمینه فعال شده قبلی در موقعیت های 2 یا 3 ریبوز نوکلئوتید tRNA باقیمانده شرکت می کند:
در این کمپلکس اسید آمینه به ریبوزوم منتقل می شود و سنتز مولکول پروتئین اتفاق می افتد. آمینواسیل-tRNA سنتتاز اختصاصی است و می تواند هر دو اسید آمینه و tRNA را تشخیص دهد. در کلینیتو، بسته به تعداد اسیدهای آمینه، کمتر از 20 سنتتاز مختلف وجود دارد.
2. tRNA، که توسط یک پیوند استری به یک اسید آمینه مرتبط است، به ریبوزوم می رسد و بر اساس نوع مکملیت بین یک سه گانه خاص از نوکلئوتیدهای mRNA، که کدون نامیده می شود و نوکلئوتیدهای سه گانه خاص مکمل آن (آنتیکودون) با mRNA تعامل می کند. tr. بنابراین، کدون mRNA نشان دهنده تثبیت خاص یک اسید آمینه در لانکوس پپتیدی در پشت آنتی کدون tRNA مربوطه است. ریبوزوم مولکولهای mRNA را میشکند و بهطور متوالی همه کدونها را میخواند و در نتیجه ترتیب توزیع همه اسیدهای آمینه را برای تحویل به محل سنتز تعیین میکند.
سنتز یک مولکول پروتئین مستقیماً از گروه آمینه قوی به گروه کربوکسیل قوی اسید آمینه ادامه می یابد. تعیین آمینو اسید COd در سنتز لنس پلی پپتیدی متیونین است که کدون توالی نوکلئوتیدی mRNA AUG است.
شروع سنتز پلی پپتید با تثبیت دو آنتی کدون tRNA در پشت کدون های mRNA آغاز می شود. این فرآیند به وضوح شامل انرژی GTP و همچنین مشارکت تعدادی از فاکتورهای پروتئینی در شروع و پپتیدیل ترانسفراز است.
با تشکر از این آنزیم، سیالیت از پیوندهای کووالانسیبه 1200 اسید آمینه / اجزا / ریبوزوم می رسد.

طرح شروع سنتز پلی پپتید
3. پس از ایجاد دی پپتید "ناپیوسته"، tRNA ریبوزوم را غیرفعال می کند و قادر به تحویل مولکول های اسید آمینه جدید است و mRNA برای سه نوکلئوتید در ریبوزوم (پلی زوم) وارد می شود. در نتیجه حرکت (جابه جایی)، کدون آزاد موقعیتی را برای شناسایی مولکول tRNA هدف اشغال می کند. همچنین در مرحله ازدیاد طول یک اسید آمینه به لنست پلی پپتیدی در زیرگونه به ترتیب کدون های مولکول mRNA اضافه می شود.
لانست پلی پپتیدی که با یک مولکول tRNA ترکیب می شود، به زیر واحد بزرگ ریبوزوم ثابت می شود. افزودن اسید آمینه اضافی پوست به لنس پلی پپتیدی به دلیل برهمکنش گروه آمینو اسید آمینه است که با tRNA و گروه کربوکسیل پپتید کمپلکس شده است.
4. خاتمه یا تکمیل سنتز مولکول پلی پپتیدی، کدون های پایانی «بدون حس» و فاکتورهای پایان پروتئین تولید می کند. سه کدون (UAG، UGA، UAA) وجود دارد که هیچ اسید آمینه ای را رمزگذاری یا متصل نمی کنند، زیرا در سلول هیچ آنتی کدون tRNA مکملی برای آنها وجود ندارد. از نظر تئوری، تنها یک کدون "بدون حس" که توسط پلی زوم در طی عبور 5-3 mRNA تولید می شود، مسئول سنتز آهسته مولکول پروتئین است.
وجود کدون پایانی در هر دنباله ای از mRNA به معنای خاتمه است سنتز پروتئین. در نتیجه، پلی زوم متلاشی می شود، mRNA نویس کریستان توسط پلی نوکلئوتید فسفوریلاز هیدرولیز می شود و زیر واحدهای ریبوزومی قبل از سنتز یک مولکول پروتئین جدید آماده می شوند.
mRNA می تواند مکرراً در فرآیند بیوسنتز پروتئین شرکت کند. عملکرد مولکول mRNA در موجودات مختلف متفاوت است. این می تواند از بسیاری از hvilins تا بسیاری از dibs متفاوت باشد.
5. DNA ساختار پروتئین اصلی را کد می کند. بنابراین، مولکول های پروتئین سنتز شده روی ریبوزوم ها هنوز در حالت نهایی خود هستند. آنها نشان دهنده پلی پپتیدهای اولیه هستند که سپس با تغییرات عددی (ارتباط مونومرها با الیگومرهای اصلاح شده، افزودن کوآنزیم ها، واکنش های شیمیایی) که ساختار پروتئین و همچنین فعالیت آنها را تغییر می دهد، شناسایی می شوند.
ساختارهای ثانویه و ثالثه کدگذاری نمی شوند، آنها با قدرت ساختار اولیه تعیین می شوند و این بدان معنی است که شکل دیگری از مولکول پروتئین در توالی اسیدهای آمینه و احتمال برهمکنش آنها با یکدیگر قرار دارد. تغییرات ساختاری پروتئین هایی که سنتز می شوند حتی روی ریبوزوم ها یا پس از اتمام سنتز در نتیجه افزودن پروتئین های مختلف انجام می شود. گروه های عاملی.
به نمودار انتقال در یک نگاه نگاه کرد
ممکن است در تناسب و شروع تغییر کنید. بنابراین، در ویروس هایی که DNA را از بین نمی برند، اطلاعات در RNA ذخیره می شود. هنگامی که ویروس به سلول نفوذ می کند، این اطلاعات به DNA سلول منتقل می شود و بقیه سپس mRNA را سنتز می کنند که پروتئین های ویروسی بر روی آن سنتز می شوند. این فرآیند رونویسی دروازه نامیده می شود و طرح انتقال در این مورد به صورت زیر خواهد بود:
در حالی که توالی نوکلئوتیدهای DNA و بنابراین mRNA حفظ می شود، ماهیت پروتئینی که به تازگی ساخته شده است بدون تغییر باقی می ماند.
اطلاعات ژنتیکی لازم برای سنتز پروتئین را می توان به روشی مشابه با ضبط زبان انسان نشان داد که از دنباله حروفی تشکیل شده است که کلمات و گفتار را تشکیل می دهد. با این حال، ماده ژنتیکی بیش از یک پایه واقعی دارد (آدنین، گوانین، اوراسیل، سیتوزین).
کد ژنتیکی شامل کلمات سه کلمه ای است. اگر در این بخش (43) قرار دهید، 64 نوع (کلمه) دریافت خواهید کرد که برای رمزگذاری 20 اسید آمینه کافی است. بنابراین، 64 کدون کد ژنتیکی را تشکیل می دهند (جدول 3).
تجزیه و تحلیل کد ژنتیکی نشان می دهد که اسیدهای آمینه مختلف تعداد کدون های متفاوتی دارند. به عنوان مثال، متیونین و تریپتوفان تنها حاوی یک کدون هستند، در حالی که آرژنین، لوسین و سرین حاوی شش کدون هستند. وجود کدون های متعدد برای یک اسید آمینه نشان دهنده «بکارت» کد است. خوب، همان اسید آمینه را می توان در پشت سه قلوهای نوکلئوتیدی خودش رمزگذاری کرد. در همان زمان، سه گانه پوستی توسط کل اسید آمینه موجود در لانست پلی پپتیدی که در حال سنتز است، شناسایی می شود.
جدول 3
کد ژنتیکی
|
نوکلئوتید |
نوکلئوتیدهای دیگر |
نوکلئوتید |
|||
کد ژنتیکی جهانی و در گونه هایی با سطوح مختلف رشد (مردم، حیوانات، گیاهان، میکروارگانیسم ها) یکسان است. جهانی بودن کد نشان می دهد که همه موجودات زنده متعلق به یک نیای واحد هستند.
به عنوان مثال، سایر اسیدهای آمینه (هیدروکسی پرولین، اکسی لیزین)، کدون را تغییر نمی دهند و با استفاده از موارد اضافی تأیید می شوند. واکنش های شیمیاییدر حال حاضر پس از سنتز لانست پلی پپتیدی. این فرآیند اصلاح پس از ترجمه نامیده می شود و برای عملکرد مناسب پروتئین پوست بسیار مهم است.
کدون های غیرگلوتینی (UAA، UAG، UGA) آمینو اسیدها را رمزگذاری نمی کنند، اما در واقع نشان دهنده اتمام سنتز یک مولکول پروتئین هستند.
بنابراین، mRNA حامل مستقیم اطلاعات ژنتیکی از هسته به ریبوزوم در سیتوپلاسم است. یک ریبوزوم تقریباً 80 نوکلئوتید در هر مکان mRNA را اشغال می کند و قادر است تقریباً 100 پیوند پپتیدی را در هر خط کاتالیز کند (Severin E. S. et al., 2011).
مولکول های پروتئین سنتز شده می توانند همزمان با ریبوزوم ها یا پس از اتمام سنتز در نتیجه افزودن گروه های عاملی مختلف، در معرض تغییرات ساختاری قرار گیرند. سیتوپلاسم mRNA دوره نسبتا کوتاهی دارد. مقدار زیادی mRNA به شکل غیرفعال سنتز و ذخیره می شود و آماده سنتز سریع پروتئین است. بسیاری از اطلاعات موجود در mRNA به یک توالی خطی از نوکلئوتیدها مرتبط است و یکپارچگی این توالی بسیار مهم است. ضایعات یا تغییر در ترتیب نوکلئوتیدها می تواند سنتز پروتئین را تغییر دهد. تا به امروز، تعدادی از بازدارنده های کم همانندسازی DNA در سلول ها (آنتی بیوتیک ها، عوامل شیمیایی، داروهای ضد ویروسی) ایجاد شده است. تغییر در توالی بازهای پورین یا پیریمیدین در یک ژن، جهش نامیده می شود.
جایگزینی فقط یک نوکلئوتید در کدون (جهش) منجر به تغییر کدگذاری یک اسید آمینه به اسید آمینه دیگر می شود. به عنوان مثال، جهش با جایگزینی اسید گلوتامیک با والین در مولکول هموگلوبین همراه است که منجر به سنتز هموگلوبین می شود که باعث کم خونی داسی شکل می شود. امروزه بیش از 200 جهش در هسته پلی پپتیدی مولکول هموگلوبین انسانی شناخته شده است. اغلب جهشزاها ترکیباتی هستند (مثلاً جایگزینهای نیتروژن) که ساختار بازهای نیتروژنی را تغییر میدهند و منجر به تغییر در ماهیت مکملی بازها میشوند. اشعه ماوراء بنفش با تشکیل دیمرهای تیمین باعث تراکم تیمین اضافی می شود. خوشبختانه به دلیل هجوم ناگوار اشعه ماوراء بنفش، این موجودات توسط توپ از جو اوزون محافظت شدند.
آنتی بیوتیک های زیادی وجود دارند که در دامپزشکی برای مهار سنتز پروتئین باکتریایی (لینکومایسین، اریترومایسین، کلرامفنیکل) حتی در مرحله ترجمه استفاده می شوند. هنگامی که این اتفاق می افتد، بافت میکروبی ژن شروع به کند کردن رشد خود می کند. آنتی بیوتیک هایی مانند تتراسایکلین ها بر سنتز ریبوزومی در سلول های موجودات زنده تأثیر نمی گذارند. پنی سیلین ها بازدارنده مستقیم سنتز پروتئین نیستند، اثرات آنها بر روی باکتری ها با مسدود کردن سنتز هگزاپپتیدهای دیواره سلولی مرتبط است. لازم به ذکر است که سنتز پروتئین نه تنها روی ریبوزوم ها، بلکه در میتوکندری ها نیز اتفاق می افتد. میتوکندری یک مکانیسم اضافی و مستقل برای سنتز پروتئین برای نیازهای خود فراهم می کند، اگرچه همه پروتئین های میتوکندری در این اندامک ها سنتز نمی شوند. RNA میتوکندری کمتر از 3٪ از محتوای RNA سلول می شود. ریبوزوم های میتوکندری از نظر اندازه کوچکتر و سیتوپلاسمی کمتری دارند. کدون UGA، به عنوان یک پایان دهنده برای سنتز پروتئین در سیتوپلاسم، در میتوکندری در کنار کدون UGG برای رمزگذاری اسید آمینه سنتز می شود.
پروتئین های سنتز شده روی ریبوزوم ها حالت کاملاً کاملی از خود باقی نمی گذارند. آنها پلی پپتیدهای اولیه را نشان می دهند که سپس با تغییرات عددی (ارتباط مونومرها با الیگومرها، افزودن کوآنزیم ها، واکنش های شیمیایی) که ساختار پروتئین و، خوب، فعالیت یوگو را تغییر می دهند، شناسایی می شوند.
تصویر 9 از ارائه "بیوسنتز پروتئین"قبل از درس زیست شناسی با موضوع "بیوسنتز پروتئین"
ابعاد: 960 x 720 پیکسل، فرمت: jpg. برای دانلود ایمن یک عکس درس زیست شناسی، روی تصویر کلیک راست کرده و روی "ذخیره تصاویر به عنوان..." کلیک کنید. برای نمایش تصاویر در درس، همچنین می توانید ارائه "Protein Biosynthesis.pptx" را به همراه تمام تصاویر در یک آرشیو فشرده به صورت رایگان دانلود کنید. حجم آرشیو 1719 کیلوبایت است.
ارائه خود را درگیر کنید"عملکرد پروتئین ها" - به این ترتیب سیگنال ها از محیط خارجی دریافت می شود و اطلاعات به سلول منتقل می شود. هنگامی که 1 گرم پروتئین به محصولات نهایی تجزیه می شود، 17.6 کیلوژول قابل مشاهده است. renaturation چیست؟ نوع کیسه: 9. کاتالیزوری. فرآیند به روز رسانی ساختار پروتئین پس از دناتوراسیون را renaturation می گویند. پیمنوف A.V. پروتئین ها یکی از منابع انرژی در بدن هستند.
"Proteins rechovina" - به عنوان مثال: کلاژن. نویسنده زیست شناسی: Boldireva L. A. ما 20 AK را می بینیم که حاوی پروتئین هستند. . باسن: تخم مرغ آب پز. آمینو اسید - گفتار ارگانیکپروتئین های غیر ارگانیک فیبریلی هستند. پروتئین های خارچوی. . سفیده های خشک ساختار پروتئین ارگانیسم Vikoristovuyutsya برای روحو. پروتئین های انرژی
"پروتئین ها و عملکرد آنها" - نقش کاتالیزوری. عملکرد موتور. مفهوم سنجاب. هیدرولیز پروتئین تا زمانی که پیوندهای پلی پپتیدی شکسته شود ادامه می یابد: خلاصه: پروتئین ها رگ های خونی، تاندون ها و مو را تولید می کنند. وظایف پروتئین چیست؟ قدرت شیمیاییپروتئین پروتئین ها در تشکیل غشای سلولی، ارگانوئیدها و غشای سلولی شرکت می کنند.
"بیوسنتز پروتئین" - فهرست منابع. ورود 4. Zmіst. بیوسنتز پروتئین ها در سلول های زنده 7. 10. 9. طرح گلاب و کلیتین پخته شده. 5. 6. 1. 8. 2. 3.
"بیوسنتز پروتئین ها" - ترجمه (لاتین: انتقال، ترجمه). رونویسی (لات. بازنویسی). با خودت چک کن اهمیت پروتئین ها Zmіst. انرژی بیوسنتز نقش آنزیم ها سنتز لنس پلی پپتیدی روی ریبوزوم ها 5. توالی نوکلئوتیدهای i-RNA ثبت شده روی قطعه DNA چیست: T-A-C-G-G-A-T-C-A-C-G-A -G-T-G-C-T A-U -G-C-G-U-A-G-U-G-C-U A-U-G-C-C-U-A-T-C-A-C-G-A -G-T-G-C-T A-U-G-C-G-U-A-G-U-G-C-U A-U-G-C-C-U-A-C-A-C-G-A
"بیوسنتز زیست شناسی پروتئین" - میکولا کوستیانتینوویچ کولتسف (1872-1940). A.G. عملکرد اصلی ریبوزوم ها سنتز پروتئین است. دگم اصلی (مقول اساسی) زیست شناسی مولکولی سنتز ماتریس است. ج. آنتی کدون - سه گانه نوکلئوتید در بالای tRNA. بیوسنتز پروتئین. پس از اتمام سنتز، iRNA به نوکلئوتیدها تجزیه می شود.
این موضوع در مجموع 8 ارائه دارد
© A.S. اسپرین
بیوسنتز پروتئین ها، SVIT RNAمانند. اسپرین
اسپرین الکساندر سرگیویچ- آکادمیک، مدیر موسسه پروتئین آکادمی علوم روسیه، عضو هیئت رئیسه آکادمی علوم روسیه.
در همان زمان، در سال 1953، D. Watson و F. Crick اصل سازماندهی ساختاری (مولکولی) مولکول ژن - اسید دئوکسی ریبونوکلئیک (DNA) را کشف کردند. ساختار DNA کلید مکانیسم ایجاد دقیق - تکرار مجدد - بیان ژن را فراهم کرد. این گونه بود که علم جدید زیست شناسی مولکولی شناخته شد. به اصطلاح جزم مرکزی زیست شناسی مولکولی فرموله شد: DNA و RNA و پروتئین ها. مفهوم در این واقعیت نهفته است که اطلاعات ژنتیکی، ثبت شده در DNA، به شکل پروتئین، اما نه به طور مستقیم، بلکه از طریق یک پلیمر اضافی - اسید ریبونوکلئیک (RNA) محقق می شود و این منجر به اسیدهای نوکلئیک اسید به پروتئین غیرقابل مذاکره می شود. . به این ترتیب، DNA بر روی DNA سنتز می شود و قدرت تکرار را تضمین می کند، به طوری که ماده ژنتیکی خروجی در نسل ها ایجاد می شود. RNA بر روی DNA سنتز می شود و در نتیجه بازنویسی و رونویسی اطلاعات ژنتیکی در قالب کپی های عددی RNA صورت می گیرد. مولکول های RNA الگوهایی برای سنتز پروتئین هستند - اطلاعات ژنتیکی به شکل لنس های پلی پپتیدی ترجمه می شود. در انواع خاص، RNA را می توان به شکل DNA ("رونویسی بازگشتی") و همچنین به شکل RNA (تکثیر) کپی کرد، اما پروتئین نمی تواند به عنوان الگوی اسیدهای نوکلئیک عمل کند (گزارش).
علاوه بر این، DNA خود نشان دهنده سیال بودن موجودات، مجموعه پروتئین هایی است که در نسل ها ایجاد می شود و علائم مرتبط با آنها. بیوسنتز پروتئین فرآیند مرکزی ماده زنده است و اسیدهای نوکلئیک از یک سو برنامه ای را در اختیار آن قرار می دهند که به معنای کل مجموعه و ویژگی پروتئین های سنتز شده است و از سوی دیگر مکانیسم ایجاد دقیق این برنامه در نسل ها بنابراین، زندگی در شکل سلولی فعلی خود به مکانیسم کاهش بیوسنتز پروتئین کاهش می یابد.
بیوسنتز پروتئین ها
دگم اصلی زیست شناسی مولکولی انتقال اطلاعات ژنتیکی از اسیدهای نوکلئیک به پروتئین ها و بنابراین به اجزای یک موجود زنده را فرض می کند. توسعه مکانیسمهایی برای اجرای این فرآیند در یک دوره ده ساله، که به دنبال فرمولبندی جزم مرکزی، عملکردهای بسیار متنوع RNA را آشکار کرد، نه تنها به عنوان یک حامل اطلاعات از ژنها (DNA) به پروتئینها و خدمت به آن. به عنوان ماتریکس برای سنتز پروتئین.
در شکل شکل 1 یک نمودار اساسی از بیوسنتز پروتئین در سلول ها را نشان می دهد. RNA پیام رسان(RNA پیام رسان، RNA پیام رسان، mRNA)، که پروتئین ها را کد می کند، همانطور که در بالا ذکر شد، تنها یکی از سه کلاس اصلی RNA های سلولی است. جرم اصلی (حدود 80٪) به کلاس دیگری از RNA تبدیل می شود - RNA ریبوزومی،که چارچوب ساختاری و مراکز عملکردی ذرات جهانی سنتز پروتئین - ریبوزوم ها را ایجاد می کند. RNA ریبوزومی خود - هم از نظر ساختاری و هم از نظر عملکرد - در تشکیل ماشینهای مولکولی فوقمیکروسکوپی به نام ریبوزوم منحصربهفرد است. ریبوزومها اطلاعات ژنتیکی را بهعنوان مولکولهای mRNA دریافت میکنند و با برنامهریزی بقیه، پروتئینهایی تولید میکنند که دقیقاً شبیه این برنامه هستند.
پروتئین، به منظور سنتز پروتئین ها، بدون اطلاعات یا برنامه کافی نیست - مواد مورد نیاز که از آنها می توان استفاده کرد. جریان مواد برای سنتز پروتئین در ریبوزوم ها با کمک دسته سوم RNA های سلولی اتفاق می افتد. حامل های RNA(RNA انتقالی، RNA انتقالی، tRNA). آنها اسیدهای آمینه را که ماده ساختمانی پروتئین ها هستند و به شکل آمینواسیل-tRNA در ریبوزوم یافت می شوند، به صورت کووالانسی متصل می کنند - می پذیرند. در ریبوزومها، aminoacyl-tRNA با کدونها - ترکیبهای trinucleotide - mRNA برهمکنش میکند و در نتیجه کدونها را در طول فرآیند ترجمه رمزگشایی میکند.
اسیدهای ریبونوکلئیک
بنابراین، ما می توانیم RNA های سلولی اصلی، فرآیند اصلی اولیه ماده زنده روزانه - بیوسنتز پروتئین را به خدمت بگیریم. Ce mRNA، RNA ریبوزومی و tRNA. RNA با کمک آنزیم ها - RNA پلیمرازها که رونویسی را انجام می دهند - روی DNA سنتز می شود - بازنویسی بخش های منفرد (واحدهای خطی) DNA دو رشته ای به شکل RNA تک رشته ای. بخشهایی از DNA که پروتئینهای سلولی را رمزگذاری میکنند، به شکل mRNA بازنویسی میشوند، به طوری که برای سنتز نسخههای متعدد RNA و tRNA ریبوزومی، بخشهای خاصی از ژنوم سلول وجود دارد که بدون ترجمه و پروتئین بیشتر، بازنویسی فشردهای را پشت سر میگذارند.
ساختار شیمیایی RNA
از نظر شیمیایی، RNA بسیار شبیه DNA است. گفتار توهین آمیز یک پلیمر خطی کامل از نوکلئوتیدها است. مونومر پوست - نوکلئوتید - یک N-گلیکوزید فسفریله شده است که در اثر بیش از حد کربن پنج کربنی - پنتوز ایجاد می شود که حامل یک گروه فسفات بر روی گروه هیدروکسیل پنجمین اتم کربن (پیوند کربن تاشو) و یک پایه نیتروژن در اولین اتم کربن (N-گلیکوزین) تفاوت شیمیایی اصلی بین DNA و RNA در این است که مونومر RNA دارای سریبوز است و مونومر DNA دارای دئوکسی ریبوز است که شبیه ریبوز است که دارای یک گروه هیدروکسیل در اتم کربن دیگر i است (شکل 1). 2).
 کم اهمیت 2.فرمول های شیمیایی مازاد یکی از ریبونوکلئوتیدها - uridylic اسید (U) و همولوگ به yomu دئوکسی ریبونوکلئوتید - اسید تیمیدیلیک (dT) |
دو نوع باز نیتروژنی در DNA و RNA وجود دارد: دو پورین - آدنین (A) و گوانین (G) - و دو پیریمیدین - سیتوزین (C) و اوراسیل (U) یا تیمین متیله (T). اوراسیل مشخصه مونومرهای RNA و تیمین مشخصه مونومرهای DNA است و این نوع دیگری از RNA و DNA است. مونومرها - ریبونوکلئوتیدهای RNA یا دئوکسی ریبونوکلئوتیدهای DNA - از طریق تشکیل اضافی محل های فسفودی استر بین اضافی های خونی (بین اتم های پنجم و سوم کربن پنتوکربن) یک نیزه پلیمری را تشکیل می دهند. بنابراین، لانست پلیمری اسید نوکلئیک - DNA یا RNA - را می توان به عنوان یک ستون فقرات قند-فسفات خطی با بازهای نیتروژنی به عنوان گروه های بیولوژیکی نشان داد. ساختار ماکرومولکولی RNA شباهت درشت ساختاری اساسی دو نوع اسید نوکلئیک در این واقعیت نهفته است که DNA یک مارپیچ فرعی منفرد است، یک درشت مولکول از دو رشته پلیمری بافته شده مکمل، که به صورت مارپیچی حول محور جانبی پیچ خورده است (بخش [ , ]) و RNA یک پلیمر تک رشته ای در عین حال، برهمکنش گروه های بیولوژیکی - پایه های نیتروژنی - یکی با یک، و همچنین با فسفات ها و هیدروکسیل های ستون فقرات قند-فسفات، منجر به این واقعیت می شود که یک پلیمر RNA تک رشته ای روی خود می سوزد و به یک می پیچد. ساختار فشرده، شبیه به احتراق یک پروتئین پلی پپتیدی nsyuga در یک کروی فشرده بنابراین، توالی های نوکلئوتیدی منحصر به فرد RNA می توانند ساختارهای فضایی منحصر به فردی را تشکیل دهند. |
برای اولین بار، ساختار فضایی خاص RNA زمانی که ساختار اتمی یکی از tRNA ها در سال 1974 رمزگشایی شد نشان داده شد. [،] (شکل 3). حنجره tRNA نیزه پلیمری که از 76 مونومر نوکلئوتیدی تشکیل شده است برای تشکیل یک هسته کروی بسیار فشرده تشکیل شده است که از آن دو برآمدگی مستقیماً به هم مالیده می شوند. آنها مارپیچ های زیر رشته ای کوتاهی روی هسته DNA هستند، اما از طریق برهمکنش بخش هایی از همان رشته RNA سازماندهی می شوند. یکی از آنها به عنوان پذیرنده اسید آمینه عمل می کند و در سنتز پروتئین پلی پپتیدی روی ریبوزوم شرکت می کند و دیگری برای برهمکنش مکمل با سه گانه (کدون) mRNA که در همان ریبوزوم کد می کند، استفاده می شود. فقط این ساختار به گونهای طراحی شده است که به طور خاص با پروتئین-آنزیمی که اسید آمینه را به tRNA متصل میکند و ریبوزوم در حین ترجمه تعامل داشته باشد، به طوری که به طور خاص توسط آنها "شناسایی" شود.

کم اهمیت 3.مدل های اتمی (چپ دست) و اسکلتی (راست دست) tRNA فنیل آلانین مخمر
تزریق RNA های ریبوزومی جدا شده راه حلی پیشرفته برای تشکیل ساختارهای فشرده و خاص از پلیمرهای خطی بلندتر از این نوع ارائه کرده است. ریبوزوم از دو بخش ناهموار تشکیل شده است - زیر واحدهای ریبوزومی بزرگ و کوچک (زیر واحد). زیر ذره پوست از یک RNA با پلیمر بالا و تعدادی پروتئین ریبوزومی مختلف تشکیل شده است. تعداد RNA های ریبوزومی حتی قابل توجه است: به عنوان مثال، RNA زیر واحد کوچک ریبوزوم باکتری حاوی بیش از 1500 نوکلئوتید است و RNA زیر واحد بزرگ تقریباً شامل 3000 نوکلئوتید است. در انسان، از جمله انسان، ciRNA حتی بزرگتر است - حدود 1900 نوکلئوتید و بیش از 5000 نوکلئوتید در زیر واحدهای کوچک و بزرگ گونه.
نشان داده شده است که RNA های ریبوزومی جدا شده از شرکای پروتئینی خود افزایش یافته و از نگاه ناب، ساختارها خود به خود به یک ساختار فشرده فرو می ریزند که از نظر اندازه و شکل مشابه زیر واحدهای ریبوزومی است]. شکل زیر ذرات بزرگ و کوچک متفاوت است و شکل RNA ریبوزومی بزرگ و کوچک بر این اساس متفاوت است (شکل 4). بنابراین، لانستهای خطی RNA ریبوزومی در ساختارهای فضایی خاص خود سازماندهی میشوند، که اندازه، شکل و احتمالاً مکان داخلی زیرواحدهای ریبوزومی و در نتیجه کل ریبوزوم را تعیین میکنند.
RNA جزئی در دنیا، تلقیح اجزای سلولهای زنده و سایر بخشهای RNA سلول کل نشان داده است که سه نوع سر RNA سمت راست قابل جداسازی نیستند. معلوم شد که در طبیعت هیچ نوع دیگری از RNA وجود ندارد. این همان چیزی است که ما آن را "RNA کوچک" می نامیم، که حاوی حداکثر 300 نوکلئوتید است، اغلب با عملکردهای ناشناخته. به عنوان یک قاعده، آنها با یک یا چند پروتئین مرتبط هستند و در سلول ها به عنوان ریبونوکلئوپروتئین - "RNP های کوچک" نشان داده می شوند.
RNA های کوچک در تمام قسمت های سلول از جمله سیتوپلاسم، هسته، هسته و میتوکندری وجود دارند. اکثر این RNP های کوچک، که عملکردهای آنها در مکانیسم های پردازش پس از رونویسی انواع اصلی RNA (پردازش RNA) - تبدیل پیش سازهای mRNA بر روی mRNA بالغ (پیچیدن)، mRNA ویرایش شده، بیوژنز tRNA، دوز ریبوزومی ایوانی شرکت می کند. RNA ها یکی از فراوانترین انواع RNPهای کوچک (SRP) در سلولها، نقش کلیدی در انتقال پروتئینهایی دارد که در طول غشای سلولی سنتز میشوند. انواع RNA های کوچکی وجود دارند که عملکردهای تنظیمی را در ترجمه انجام می دهند. یک RNA کوچک ویژه وارد انبار مهم ترین آنزیم مسئول پشتیبانی از تکثیر DNA در نسل های سلولی - تلومراز می شود. باید گفت که ابعاد مولکولی آنها می تواند برابر با پروتئین های کروی سلولی باشد. بنابراین، به طور فزاینده ای روشن می شود که عملکرد یک سلول زنده نه تنها با تنوع پروتئین هایی که در آن سنتز می شوند، بلکه با وجود مجموعه ای غنی از RNA های متنوع، از جمله RNA های کوچک، تعیین می شود. فشردگی و اندازه سلول پروتئین ها قابل توجه است.
ریبوزیم. تمام زندگی فعال توسط تبادل مواد - متابولیسم هدایت می شود، و تمام واکنش های بیوشیمیایی متابولیسم با مایعات لازم برای اطمینان از زندگی انجام می شود، فقط با کاتالیزورهای خاص بسیار موثر، اجازه دهید تکامل را ایجاد کنیم. برای چندین دهه، بیوشیمی متقاعد شده است که کاتالیز بیولوژیکی همیشه توسط پروتئین هایی به نام انجام می شود. آنزیم ها، یا آنزیم ها I axis y 1982-1983 pp. نشان داده شده است که در طبیعت انواعی از RNA وجود دارد که مانند پروتئین ها دارای فعالیت کاتالیزوری بسیار ویژه هستند [،]. این کاتالیزورهای RNA نامگذاری شدند ریبوزیم هادانش در مورد گناه پروتئین ها در کاتالیز واکنش های بیوشیمیایی به پایان رسیده است.
امروزه ریبوزوم را ریبوزیم نیز می دانند. درست است، تمام داده های تجربی آشکار نشان می دهد که سنتز پروتئین پلی پپتیدی در ریبوزوم توسط RNA ریبوزومی کاتالیز می شود و نه توسط پروتئین های ریبوزومی. یک جزء کاتالیزوری از RNA ریبوزومی بزرگ شناسایی شده است که مسئول کاتالیز واکنش transpeptidation است که همچنین به رشد پلی پپتید پروتئین در طول ترجمه کمک می کند.
در مورد تکثیر DNA ویروسی، مکانیسم آن تفاوت کمی با همانندسازی مواد ژنتیکی - DNA - خود سلول دارد. در برخی از RNA های ویروسی، فرآیندهایی انجام می شود که سرکوب می شوند یا در تمام طول روز در سلول های طبیعی، که در آن تمام RNA تنها بر روی DNA مانند یک ماتریکس سنتز می شود. هنگامی که با ویروس های RNA آلوده می شود، وضعیت می تواند دوگانه باشد. در برخی موارد، DNA بر روی RNA ویروسی سنتز می شود، مانند یک ماتریکس ("رونویسی بازگشتی")، و روی این DNA نسخه های متعددی از RNA ویروسی رونویسی می شود. در موارد دیگر، که به ما مربوط است، یک رشته مکمل از RNA بر روی RNA ویروسی سنتز می شود که به عنوان الگویی برای سنتز - همانندسازی - نسخه های جدید RNA ویروسی عمل می کند. به این ترتیب، در هنگام عفونت با ویروس های RNA، اصل تعیین کننده RNA که ایجاد ساختار آن را تعیین می کند، همانطور که DNA نیز محقق می شود.
چند کارکردی RNA فرضیه و دانش کلی در مورد عملکرد RNA به ما اجازه می دهد تا در مورد عملکرد منحصر به فرد این پلیمر در طبیعت زنده صحبت کنیم. امکان ایجاد تغییر فوری در عملکردهای اساسی RNA وجود دارد.
عملکرد همانندسازی ژنتیکی: توانایی ساختاری کپی (تکثیر) توالی های نوکلئوتیدی خطی از طریق توالی های مکمل. این عملکرد در طول عفونت های ویروسی اجرا می شود و شبیه عملکرد اصلی DNA در موجودات زنده سلولی - تکرار مجدد مواد ژنتیکی است.به طور کلی، RNA بهعنوان یک پلیمر شگفتانگیز در برابر ما قرار دارد، که به نظر میرسد هیچ زمانی برای تکامل کیهان، و نه عقل خالق اثری از خود بر جای نمیگذاشت. همانطور که مشخص است، RNA برای ترکیب عملکرد هر دو پلیمر اساسی - DNA و پروتئین ها طراحی شده است. جای تعجب نیست که تغذیه مقدم بر علم شده است: چرا احساس گناه و وجود خودکفای نور RNA نمی تواند ظاهر زندگی را به شکل پروتئین DNA فعلی آن منتقل کند؟تابع کدگذاری: برنامه ریزی سنتز پروتئین توسط توالی های خطی نوکلئوتیدها. این همان عملکردی است که DNA دارد. من در DNA و RNA همان سه گانه نوکلئوتیدها 20 اسید آمینه پروتئین را کد می کنند و توالی یک سه گانه در اسید نوکلئیک برنامه ای برای آرایش متوالی 20 نوع اسید آمینه در یک لانست سفید پلی پپتیدی است.
عملکرد سازه ساز: تشکیل ساختارهای بی اهمیت منحصر به فرد. مولکولهای کوچک RNA که بهطور فشرده تا شدهاند، اساساً شبیه ساختارهای بیاهمیت پروتئینهای کروی هستند و مولکولهای RNA بزرگتر میتوانند بخشهای بیولوژیکی بزرگتر یا هستههای آنها را ایجاد کنند.
عملکرد شناسایی: برهمکنش های بسیار خاص با سایر ماکرومولکول ها (از جمله پروتئین ها و سایر RNA ها) و با لیگاندهای کوچک. این عملکرد احتمالاً اصلی ترین عملکرد در پروتئین ها است. این بر اساس توانایی پلیمر برای تشکیل یک ساختار منحصر به فرد و تشکیل ساختارهای بی اهمیت خاص است. تابع تشخیص اساس کاتالیزور خاص است.
عملکرد کاتالیزوری: کاتالیز اختصاصی واکنش های شیمیایی توسط ریبوزیم ها. این عملکرد مشابه عملکرد آنزیمی پروتئین های آنزیمی است.
LIFE HAVEN
نظریه Oparin's protein-coacervate. شاید، طبق علم، یک نظریه خوب فکر شده از زندگی در مسیر زیست زایی توسط بیوشیمیدان A.I. اوپارین در دهه 20 قرن گذشته متولد شد [،]. این تئوری مبتنی بر کشف این بود که همه چیز با پروتئین ها آغاز شد، و در ذهن های اولیه امکان سنتز شیمیایی خود به خود مونومرهای پروتئین - اسیدهای آمینه - و پلیمرهای پروتئین مانند (پلی پپتیدها) از طریق او وجود داشت. انتشار این نظریه باعث تحریک آزمایش های عددی در تعدادی از آزمایشگاه ها در سراسر جهان شد که واقعیت چنین ترکیبی را در ذهن افراد نشان داد. نظریه شویدکو به طور گسترده پذیرفته شد و بسیار محبوب شد.
فرض اصلی آن این بود که ذرات پروتئین مانند به طور خود به خود در "آبگوشت" اصلی ظاهر می شوند و "در قطره های coacervate - یک سیستم کلوئیدی تقویت شده (sols) که در یک محلول آب رقیق تر شناور می شوند، ترکیب می شوند. این باعث تغییر ذهنیت شد. گناه ارگانیسم ها - تقویت dovkillaاز آنجایی که بخشهای مختلف پروتئین مانند لکههای کواسروات میتوانستند فعالیت کاتالیزوری داشته باشند، ممکن بود واکنشهای بیوشیمیایی در سنتز در وسط لکهها رخ دهد. تجزیه بیشتر به قطعات - تولید مثل. کواسروات که جذب، رشد و تکثیر می شود و با تقسیم بندی به عنوان نمونه اولیه سلول های زنده دیده می شود (شکل 5).
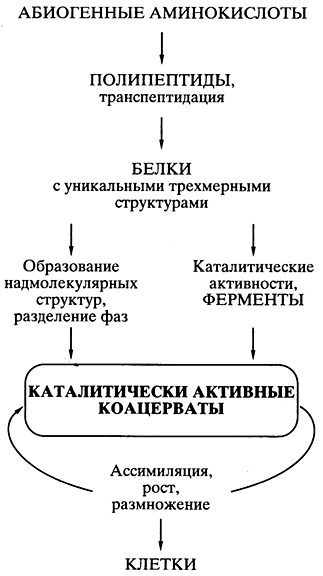
کم اهمیت 5.جملات شماتیک در مورد مسیرهای زندگی
بر اساس تئوری پروتئین کواسروات A.I. اوپارینا
همه چیز به خوبی اندیشیده شده بود و از نظر علمی مبتنی بر تئوری بود، به جز یک مشکل، که مدت هاست با چشمان همه تقلبی های زندگی گالوسا صاف شده است. از آنجایی که به طور خود به خود، در یک سری از سنتزهای تصادفی بدون الگو در یک کواسروات، ساختار مولکول های پروتئین یک به یک پدید آمدند (به عنوان مثال، کاتالیزورهای موثری که برتری این کواسروات را در رشد و تکثیر آن تضمین می کنند)، پس چگونه می توانند پلیس برای گشاد کردن وسط کواسروات و بیشتر برای انتقال به کواسروات ها استفاده می شود - نظریه مشخص شد که برجسته کردن مشکل ایجاد دقیق - در وسط یک کواسروات و در نسل ها - ساختارهای پروتئینی مؤثر منفرد بی اساس است. ناگهان ظاهر شد.
RNA سبک به عنوان جانشین زندگی روزمره. دانش انباشته در مورد کد ژنتیکی، اسیدهای نوکلئیک و بیوسنتز پروتئین منجر به تثبیت یک ایده اساساً جدید در مورد TOM شد، که همه چیز نه با پروتئین، بلکه با RNA [-] آغاز شد. اسیدهای نوکلئیک نوع واحدی از پلیمرهای بیولوژیکی هستند که ساختار ماکرومولکولی آن ها به دلیل اصل مکمل بودن در سنتز لانکساگ های جدید (گزارش ها)، امکان کپی برداری از یک توالی خطی مرطوب و نوارهای مونومر را تضمین می کند، به عبارت دیگر، امکان ایجاد (تکثیر) پلیمر، ریزساختار آن. بنابراین، تنها اسیدهای نوکلئیک، به جای پروتئین، می توانند ماده ژنتیکی باشند و مولکول هایی را ایجاد کنند که ریزساختار خاص خود را در نسل ها تکرار می کنند.
در سطوح پایین، خود RNA، و نه DNA، می تواند ماده ژنتیکی اولیه باشد.
طبق اولیو در سنتز شیمیایی و واکنش های بیوشیمیایی، ریبونوکلئوتیدها به دئوکسی ریبونوکلئوتیدها منتقل می شوند. دئوکسی ریبونوکلئوتیدها محصولات اصلاح ریبونوکلئوتیدها هستند (شکل 2).
به شکلی متفاوت،در فرآیندهای مدرن و جهانی متابولیسم زنده، خود ریبونوکلئوتیدها، به جای دئوکسی ریبونوکلئوتیدها، به طور گسترده نشان داده می شوند، از جمله حامل های اصلی انرژی از نوع پلی فسفات های ریبونوکلئوزیدی (ATP).
در سوم،همانندسازی RNA می تواند بدون هیچ بخشی از DNA رخ دهد و مکانیسم تکثیر DNA در دنیای زنده شامل بخش اتصال پرایمر RNA در شروع سنتز DNA lanjug است.
در چهارمینبا همه ماتریکس و توابع ژنتیکی مشابهی که DNA و RNA ایجاد میشوند، آنها همچنین عملکرد پایینی دارند که پروتئینها را کنترل میکنند، از جمله کاتالیز واکنشهای شیمیایی. بنابراین، هر دلیلی وجود دارد که DNA را به عنوان یک افزوده تکاملی بعدی - به عنوان یک اصلاح RNA، که برای عملکرد خاص ایجاد و حفظ نسخه های منحصر به فرد ژن ها در انبار تخصص دارد، مشاهده کنیم. ژنوم کوچک بدون نقش واسطه در بیوسنتز پروتئین.
هنگامی که RNA های فعال کاتالیزوری کشف شدند، ایده تقدم RNA در زندگی روزمره به شدت تحت تأثیر قرار گرفت و این مفهوم فرموله شد. جهان خودکفا RNA،از بین بردن زندگی روزمره [،]. نمودار Mozhliv از RNA vinification در شکل 1 ارائه شده است. 6.
سنتز ابیوژنیک ریبونوکلئوتیدها و تشکیل کووالانسی آنها به الیگومرها و پلیمرهایی از نوع RNA را می توان تقریباً در همان ذهن ها و در همان شرایط شیمیایی که برای ایجاد اسیدهای آمینه و پلی پپتیدها فرض شده بود، ایجاد کرد. اخیرا O.B. Chetverin و دانشمندان (موسسه پروتئین، آکادمی علوم روسیه) به طور تجربی نشان دادند که پلی ریبونوکلئوتیدها (RNA) که در محیط آبی پایه استفاده می شوند، قبل از نوترکیبی خود به خود ایجاد می شوند، به طوری که تبادل آب راه استری کامی lantsyuga است. مبادله قطعات لانست کوتاه با قطعات بلند منجر به کاهش پلی ریبونوکلئوتیدها (RNA) می شود و چنین نوترکیبی خود با تنوع ساختاری این مولکول ها سازگار است. از جمله آنها می توان مولکول های RNA فعال کاتالیزوری را نام برد.
ظاهر نادر مولکولهای RNA منفرد، که پلیمریزاسیون ریبونوکلئوتیدها یا اتصال الیگونوکلئوتیدها را بر روی یک ماتریس RNA مکمل [ ] کاتالیز میکنند، به معنای تشکیل یک مکانیسم رپ پیوندهای RNA است. تکثیر کاتالیزورهای RNA (ریبوزیم ها) خود آسیب کمی به جمعیت های RNA خودتکثیر شونده وارد می کند. تولید نسخه های خود از RNA تکثیر شده است. تغییرات اجتناب ناپذیر در کپی (جهش) و نوترکیب در جمعیت های RNA که خود تکرار می شوند، تنوع روزافزونی را در این جهان ایجاد کرد. بنابراین، انتقال دنیای باستانی RNA - tse "دنیای بیولوژیکی خودکفا که در آن مولکول های RNA به عنوان ماده ژنتیکی و به عنوان کاتالیزورهای آنزیم مانند عمل می کنند." .
سرزنش بیوسنتز پروتئین علاوه بر این، بر اساس RNA نور، پیشرفت کمی در شکلگیری مکانیسمهای بیوسنتز پروتئین، ظهور پروتئینهای مختلف از ساختار و قدرت فروپاشیده، تقسیمبندی سیستمهای بیوسنتز پروتئین و مجموعههای پروتئین، شاید به شکل کواسرواتها وجود دارد. و تکامل ساختار بافت باقیمانده - سلولهای زنده (بخش شکل 6). ).
مسئله گذر از نور RNA باستانی به نور سنتز کننده پروتئین فعلی مهمترین نکته برای یک راه حل بسیار نظری است. امکان سنتز بیوژنیک پلی پپتیدها و ترکیبات پروتئین مانند به مشکل فعلی کمک نمی کند، زیرا هیچ مسیر خاصی قابل مشاهده نیست، چگونه این سنتز ممکن است شامل اتصال به RNA باشد و تحت کنترل ژنتیکی است. کنترل ژنتیکی سنتز پلی پپتیدها و پروتئین ها به طور مستقل از سنتز زیست زایی اولیه، به روش خاص خود، با تنظیم RNA نور موجود از قبل ایجاد می شود. ادبیات تعدادی فرضیه را در مورد شباهت مکانیسم روزانه بیوسنتز پروتئین در RNA سبک ارائه کرده است، اما شاید بتوان آنها را از نظر احتمالات فیزیکی-شیمیایی با جزئیات و پیش بینی نشده دید. من نسخه خود را از روند تکامل و تخصصی شدن RNA ارائه می کنم، که منجر به تخریب دستگاه بیوسنتز پروتئین می شود (7 کوچک)، اما تظاهر نمی کند که کامل شده است.
یک طرح فرضی برای ترکیب دو لحظه اساسی که به نظر مهم میرسند پیشنهاد شده است.
طبق اولیفرض بر این است که الیگوریبونوکلئوتیدهای سنتز شده به طور فعال از طریق مکانیسم اضافی ترانس استری غیر آنزیمی خود به خودی دوباره ترکیب می شوند که منجر به ایجاد رشته های RNA فعال و ایجاد تنوع آنها می شود. این مسیر در جمعیت الیگونوکلئوتیدها و پلی نوکلئوتیدها می تواند شامل انواع فعال کاتالیستی RNA (ریبوزیم) و سایر انواع RNA با عملکردهای تخصصی باشد (شکل 7). علاوه بر این، نوترکیب غیر آنزیمی اولیگونوکلئوتیدهایی که به طور مکمل به ماتریکس پلی نوکلئوتید متصل می شوند، می تواند اتصال متقابل (پیچیدن) قطعات مکمل ماتریکس را در یک لانست تضمین کند. به این ترتیب، به جای کاتالیز شدن توسط پلیمریزاسیون مونونوکلئوتیدها، کپی (تکثیر) اولیه RNA می تواند رخ دهد. ظاهراً، اگر مشخص شد که ریبوزیمها دارای فعالیت پلیمراز پایینی هستند، کارایی (دقت، سیالیت و بهرهوری) به مکمل کپی میشود. ماتریس ها کوچک می شوند و در حال رشد هستند.

کم اهمیت 7.طرح تکامل و تخصصی شدن مولکول های RNA
در فرآیند انتقال از دنیای باستانی RNA به به دنیای کنونی
بیوسنتز پروتئین تعیین شده ژنتیکی
دیگریک نکته مهم در نسخه من این است که اولین دستگاه بیوسنتز پروتئین با هماهنگی بسیاری از انواع RNA تخصصی قبل از ظهور دستگاه تکثیر آنزیمی (پلی مراز) ماده ژنتیکی - RNA i DNA ظاهر شد. این دستگاه اولیه شامل RNA پروریبوزومی فعال کاتالیستی است که فعالیت پپتیدیل ترانسفراز پایینی دارد. مجموعه ای از pro-tRNA ها که به طور خاص به اسیدهای آمینه یا پپتیدهای کوتاه متصل می شوند. سایر RNA های پروریبوزومی به طور همزمان با RNA کاتالیزوری پروریبوزومی، pro-mRNA و pro-tRNA برهمکنش می کنند (شکل 7). چنین سیستمی میتواند از طریق یک واکنش transpeptidation کاتالیز شده، لنجهای پلی پپتیدی را سنتز کند. در میان سایر پروتئین های فعال کاتالیزوری - آنزیم های اولیه (آنزیم ها) - پروتئین هایی ظاهر شده اند که پلیمریزاسیون نوکلئوتیدها - رپلیکازها یا NK پلیمرازها را کاتالیز می کنند.
با این حال، ممکن است که فرضیه در مورد به دنیای باستان RNA، به عنوان پیش ساز دنیای زندگی روزانه، نمی تواند شرطی شدن کافی را برای حل مشکل اصلی به دست آورد - یک توصیف علمی قابل قبول از مکانیسم انتقال از RNA و همانندسازی به بیوسنتز پروتئین. فرضیه جایگزین A.D اضافه شده و به تفصیل فکر شده است. آلتشتاین (موسسه زیست شناسی ژن آکادمی علوم روسیه)، که فرض می کند که تکثیر ماده ژنتیکی و ترجمه آن - سنتز پروتئین - به طور همزمان تکامل و تکامل یافته و مصرف می شود، با شروع از تعامل متقابل سنتز یونی الیگونوکلئوتیدها و آمینواسیل-نوکلئوتیدیل ها - انیدریدهای مخلوط قبلاً قزاق نزدیک شده است ... ( «و شهرزاد صبح گرفتار شد و اجازه تبلیغ گرفت».)
ادبیات
1. واتسون جی دی، کریک اف اچ سی.ساختار مولکولی اسیدهای نوکلئیک // طبیعت. 1953. ج 171. ص 738-740. 2. واتسون جی دی، کریک اف اچ سی.مفاهیم ژنتیکی ساختار اسید نوکلئیک دئوکسی ریبوز // Nature 1953 V. 171. P. 964-967. 3. اسپرین A.S.زیست شناسی روزانه و ایمنی بیولوژیکی // بولتن آکادمی علوم روسیه. 1997. شماره 7. 4. اسپرین A.S.در مورد ساختار ماکرومولکولی مواد شیمیایی ریبونوکلئیک طبیعی با پلیمر بالا // مجله زیست شناسی مولکولی. 1960. V. 2. P. 436-446. 5. کرن اس.اچ.، سودات اف.ال.، کویگلی جی.جی. تا درساختار سه بعدی RNA انتقال فنیل آلانین مخمر // علم. 1974. ج 185. ص 435-40. 6. روبرتاس جی.دی.، لدنر جی.ای.، فینچ جی.تی. تا درساختار tRNA فنیل آلانین مخمر با وضوح 3 A // طبیعت. 1974. V. 250. ص 546-551. 7. واسیلیف V.D.، Serdyuk I.N.، Gudkov A.T.، SPIRin A.S.خود سازماندهی RNA ریبوزومی // ساختار، عملکرد و ژنتیک ریبوزوم ها / ویرایش. Hardesty B. and Kramer G. New York: Springer-Verlag, 1986. P. 129-142. 8. Baserga S.J.، Steitz J.A.دنیای متنوع ریبونوکلئوپروتئین های کوچک // دنیای RNA / ویرایش. Gesteland R.F. اتکینز جی.اف. New York: Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1993, pp. 359-381. 9. Kruger K.، Grabowski PJ.، Zaug AJ. تا در. RNA خودجوش: برداشتن خودکار و اتوسیکلیزاسیون توالی مداخله گر RNA ریبوزومی تتراهیمنا// سلول. 1982. ج 31. ص 147-157. 10. Guerrier-Takada S.، Gardiner K.، Marsh T. و همکاران.ریبونوکلئازهای نصف RNA P زیرواحد کاتالیزوری آنزیم // سلول است. 1983. ج 35. ص 849-857. 11. Oparin A.I.مسیر زندگی. M: روباتنیک مسکو، 1924. 12. Oparin A.I.گناه زندگی زمین (3 نوع). م: شعبه آکادمی علوم SRSR، 1957. 13. ویز اس.تکامل کد ژنتیکی // کد ژنتیک. نیویورک: هارپر و رو، 1967. صفحات 179-195. 14. کریک F.H.C.منشا کد ژنتیکی // مجله زیست شناسی مولکولی. 1968. ج 38. ص 367-379. 15. Orgel L.E.تکامل دستگاه ژنتیکی // مجله زیست شناسی مولکولی. 1968. ج 38. ص 381-393. 16. گیلبرت دبلیو.جهان RNA // طبیعت. 1986. ج 319 ص 618. 17. جویس G.F.، Orgel L.E.چشم انداز درک منشاء جهان RNA // دنیای RNA / ویرایش. Gesteland R.F. اتکینز جی.اف. New York: Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1993 P 1-25. 18. Chetverina H.V.، Demidenko A.A.، Ugarov V.I.، Chetverin A.B.سازماندهی مجدد خود به خود در توالی های RNA // نامه های FEBS. 1999. ج 450. ص 89-94. 19. Bartel D.P., Szostak J.W.جداسازی ریبوزیم های جدید از مجموعه بزرگی از توالی های تصادفی // علم. 1993. ج 261. ص 1411-1418. 20. Ekland E.H.، Bartel D.P.پلیمریزاسیون RNA کاتالیز شده با RNA از تری فسفات های ویکوریستی و نوکلئوزیدی // طبیعت. 1996 V. 382. ص 373-376. 21. Orgel L.E.منشاء حیات - بررسی حقایق و گمانه زنی ها // گرایش های علوم بیوشیمی. 1998. V. 23. p. 491-495. 22. Altshtein A.D.شباهت سیستم ژنتیکی: فرضیه پروژن // زیست شناسی مولکولی. 1987. T. 21. صص 309-322.








