U svim živim ćelijama, proteini se sintetiziraju ribozomima . Ribosom je velika makromolekula sa presavijenom asimetričnom četvrtastom strukturom koja se sastoji od ribonukleinskih kiselina (ribosomalna RNK) i proteina. Za sintetizaciju proteina ribosom je opremljen sa:
1. Program koji postavlja redoslijed obrade viška aminokiselina polipeptidnog proteina.
2. Aminokiselinski materijal, iz kojeg će biti proteini.
3. Energija.
Sam ribosom ima katalitičku (enzimsku) funkciju, odgovornu za stvaranje peptidnih veza i, vjerovatno, polimerizaciju aminokiselinskih ostataka u polipeptidnim proteinima.
Program koji postavlja red viška aminokiselina u polipeptidnom lankusu proteina izlazi kao deoksiribonukleinska kiselina (DNK), odnosno genom ćelije. Sintetizirani RNK lanci su komplementarni jednom od lanaca DNK i na taj način precizno kreiraju deoksiribonukleotidnu sekvencu drugog lanca DNK u vlastitoj ribonukleotidnoj sekvenci. Proces kopiranja gena, koji se obrađuje enzimom RNA polimerazom, naziva se transkripcija. Tokom sinteze RNK, posebno u eukariotskim ćelijama, ona može pretrpjeti niz dodatnih promjena koje se nazivaju procesiranje, tokom kojih može promijeniti sadržaj nukleotidne sekvence. RNK koja se oslobađa ide dalje do ribozoma u jezgri programa, koji određuje sekvencu aminokiselina u sintetiziranom proteinu. Zove se informacijska ili "glasnička" RNK (mRNA). Dakle, sama transkripcija gena i stvaranje mRNA osigurava protok informacija od DNK do ribozoma.
Materijal klipa, koji proizvodi protein, sadrži aminokiseline. Međutim, ribosom ne apsorbuje slobodne aminokiseline.Da bi služila kao supstrat za ribozom, aminokiselina mora biti aktivirana kroz rezultujuće cepanje ATP-a i prihvaćena (kovalentno prihvaćena) od strane posebnog RNA molekula koji se naziva transfer ili transport RNK (tRNA), koja pomaže u sintezi enzima. Aminoacil-tRNA se uklanja iz ribozoma kao supstrat za sintezu proteina. Osim toga, energija hemijske veze između aminokiselinskog ostatka i tRNA se koristi za reakciju peptidne veze u ribosomu. Tako će aktivacija aminokiselina i stvaranje aminoacil-tRNA osigurati protok materijala i energije za sintezu ribosomskih proteina.
Ova tri toka (informacija, materijal i energija) konvergiraju u ribosomu. Primajući ih, ribosom vrši translaciju, odnosno translaciju, genetske informacije iz nukleotidne sekvence mRNA u aminokiselinsku sekvencu sintetizovanog polipeptidnog proteina. Kada se izrazi u molekularnim terminima, ribosom sekvencijalno skenira DNK mRNA (kolapsira nakon nje), a zatim sukcesivno selektuje aminoacil-tRNA iz sredine, usled čega se bira specifičnost aminoacilnog ostatka Postoji aminoacil- tRNA ribosoma koji je direktno određen specifičnošću kombinacije. mRNA fragment. Dakle, nastaje problem genetskog koda: koje kombinacije nukleotida su određene, odnosno šta kodira kožu sa 20 aminokiselina, od kojih će se formirati proteinski molekuli?
Protok ribozoma kroz ribozom mRNA (ili, drugim riječima, prolazak ribozoma kroz ribozom) postavlja strogi vremenski sat ulaska u ribozom različitih aminoacil-tRNA od podvrste do reda. rotacije nukleotidnih kombinacija koje kodiraju, čvorovi f mRNA. Aminoacilni ostatak sastavljene aminoacil-tRNK je odmah kovalentno vezan ribozomom za rastuću polipeptidnu koplju. Deacilirana tRNA se veže za ribozome kod različitih vrsta. Tako će se, uzastopno, korak po korak, formirati polipeptidni lancet proteina (div. šema 1).
Biosinteza proteina.
Plastični metabolizam (asimilacija i anabolizam) je ukupnost reakcija biološke sinteze. Ova vrsta razmjene nazvana je po svojoj suštini: iz govora, poput govora klijenta, iz poze nastaju govori, slični govorima klijenta.
Pogledajmo jedan od najvažnijih oblika plastičnog metabolizma - biosintezu proteina. Biosinteza proteina Javlja se u svim ćelijama oko eukariota. Informacija o primarnoj strukturi (poretku aminokiselina) proteinskog molekula je kodirana nizom nukleotida u produžetku molekula DNK - genima.
Gen je dio molekule DNK, što znači redoslijed aminokiselina u molekulu proteina. Takođe, nukleotidi u genu određuju redosled aminokiselina u polipeptidu. Ovo je primarna struktura u kojoj leže sve druge strukture, moći i funkcije proteinske molekule.
Sistem za snimanje genetskih informacija u DNK (i RNK) prema tačnom nizu nukleotida naziva se genetski kod. Tobto. Jedinica genetskog koda (kodon) je triplet nukleotida u DNK ili RNK koji kodira jednu aminokiselinu.
Cijeli genetski kod uključuje 64 kodona, od kojih je 61 kodirajući, a 3 nekodirajući (terminatorski kodoni, koji ukazuju na završetak procesa prevođenja).
Kodon-terminatori i - RNA: UAA, UAG, UGA, u DNK: ATT, ATC, ACC.
Početak procesa translacije je inicijatorski kodon (AUG, DNK - TAC), koji kodira aminokiselinu metionin. Ovaj kodon je prvi koji ulazi u ribozom. Zatim se metionin, budući da se ne prenosi kao prva aminokiselina ovog proteina, odvaja.
Genetski kod je karakteristika moći.
1. Univerzalnost – kod je isti za sve organizme. Isti triplet (kodon) u bilo kojem organizmu kodira istu aminokiselinu.
2. Specifičnost - svaki kodon kodira samo jednu aminokiselinu.
3. Generičnost – većina aminokiselina može biti kodirana sa nekoliko kodona. Za to su krive dvije aminokiseline - metionin i triptofan, koji su povezani samo sa jednom varijantom kodona.
4. Između gena postoje “znakovi podjele” - tri posebna tripleta (UAA, UAG, UGA), od kojih svaki ukazuje na sintezu polipeptidnog lanjuga.
5. U sredini gena nema "divizionih znakova".
Da bi se protein sintetizirao, informacije o sekvenci nukleotida u njegovoj primarnoj strukturi mogu se dostaviti ribosomima. Ovaj proces uključuje dvije faze – transkripciju i prijevod.
Transkripcija(prepisivanjem) informacije se dobijaju sintezom molekula RNK sa jednim lancem na jednoj strani molekule DNK, čiji se redosled nukleotida tačno poklapa sa sekvencom nukleotida matriksa - polinukleotidnom DNK molekulom.
Vaughn (i - RNA) je posrednik koji prenosi informacije od DNK do mjesta gdje se molekuli proteina sklapaju u ribosomu. Sinteza RNK (transkripcija) se očekuje u ranoj fazi. Enzim (RNA polimeraza) razdvaja lanac DNK, a jedan od lanaca (kodira) proizvodi RNA nukleotide na osnovu principa komplementarnosti. Molekul RNK se sintetiše na ovaj način (template synthesis) i ulazi u citoplazmu, a male podjedinice ribozoma su nanizane na jednom kraju.
Druga faza u biosintezi proteina je emitovanje- ovo je translacija sekvence nukleotida u molekulu i - RNA je sekvenca aminokiselina u polipeptidu. Kod prokariota, koji nemaju formirano jezgro, ribozomi se mogu vezati za novostvorenu molekulu i RNA enzim nakon što se odvoje od DNK ili dovesti do potpunog završetka sinteze. Kod eukariota, i-RNA se isporučuje kroz nuklearni omotač u citoplazmu. Prijenos se vrši pomoću posebnih proteina koji stvaraju kompleks s molekulom i RNK. Osim funkcije prijenosa ovih proteina, oni oduzimaju i-RNA iz djelovanja, što uništava citoplazmatske enzime.
U citoplazmu, na jednom kraju RNK (i na samom kraju gdje počinje sinteza molekula u jezgri), ulazi ribosom i počinje sinteza polipeptida. Kada je ribosom napunjen molekulom RNK, on prevodi triplet za tripletom, sukcesivno dodajući aminokiseline na zreli kraj polipeptidne petlje. Tačan identitet aminokiselinskog koda za triplet i RNK osigurava tRNA.
Transfer RNA (tRNA) "donosi" aminokiseline u veliku podjedinicu ribosoma. Molekul t-RNA ima presavijenu konfiguraciju. Na ovim plohama formiraju se vodene veze između komplementarnih nukleotida, a oblik molekula podsjeća na stabilan list. Na vrhu ekspanzije nalazi se triplet jakih nukleotida (antikodon), koji predstavlja glavnu aminokiselinu, a baza je mjesto gdje je vezana ova aminokiselina (slika 1).
Mala 1. Šema budućeg prijenosa RNK: 1 – vodene veze; 2 – antikodon; 3. mjesto vezivanja aminokiselina.
Kožna t-RNA može prenijeti vlastitu aminokiselinu. T-RNA se aktivira posebnim enzimima, prihvata svoju aminokiselinu i transportuje je do ribosoma. U sredini ribozoma trenutno postoje samo dva kodona i-RNA. Pošto je antikodon t-RNA komplementaran kodonu i-RNA, t-RNA se odmah dodaje aminokiselini i-RNA. Prije drugog kodona dodaje se još jedna t-RNA koja nosi vlastitu aminokiselinu. Aminokiseline sintetizira velika podjedinica ribosoma, a uz pomoć enzima između njih se uspostavlja peptidna veza. Istovremeno se formiraju veze između prve amino kiseline i t-RNA, te t-RNA sa ribosomima iza prve amino kiseline. Ribosom se pomiče jednu po jednu trojku i proces se ponavlja. Na taj način postepeno se uzgaja polipeptidna molekula u kojoj se formiraju aminokiseline istog tipa sa redoslijedom tripleta koji ih kodiraju (template synthesis) (slika 2).

Mala 2. Šema bisinteze proteina: 1 – i-RNA; 2 - ribosomske podjedinice; 3 – t-RNA sa aminokiselinama; 4 – t-RNA bez aminokiselina; 5 – polipeptid; 6 - kodon ta-RNA; 7-antikodon tRNA.
Jedan ribosom je sposoban da sintetiše novi polipeptidni lancet. Prote, često jedan i-RNA molekul kolabira gomilu ribozoma. Takvi kompleksi se nazivaju poliribozomi. Nakon završetka sinteze, polipeptidno lance formira se u matriks - i-RNA molekul, savijen u spiralu i formira sekundarnu (sekundarnu, tercijarnu ili kvaternarnu) strukturu. Ribosomi rade vrlo efikasno: u roku od jedne sekunde, bakterijski ribosom probavlja polipeptidni lancet koji sadrži 20 aminokiselina.
Biosinteza proteina (polipeptida) je izuzetno složen i nevjerovatan proces. Biosinteza proteina se aktivno odvija u svim organima i tkivima, uključujući eritrocite. Mnoge ćelije sintetiziraju proteine za “izvoz” (ćelije jetre, podpužve), au ovom slučaju sadrže čak i veliki broj ribozoma. U životinjskoj kulturi broj ribozoma doseže 10 5; promjer ribozoma doseže 20 nm.
Proces sinteze proteina odvija se u sredini ćelija na površini ribozoma, koji su kompleksi dve podjedinice sa konstantom sedimentacije 60S i 40S, koje funkcionišu kao jedna celina. Proteini ribosoma postaju 30-35%, a ribosomska RNK 65-70%. Ribozomi su podijeljeni na aminoacilne i peptidilne dijelove. Prvi služi za fiksaciju kompleksa aktivne aminokiseline i tRNA koji se nalazi na ribosomu, a drugi fiksira polipeptidni lancet vezan za drugu tRNA. Podjedinice ribozoma se sintetiziraju u jezgru na DNK šablonu.
Suština procesa sinteze proteina predstavljena je dijagramom:
Sistem za sintezu proteina uključuje ribozome, nukleinske kiseline, set od 20 aminokiselina, različite enzime, ATP, GTP, jone magnezijuma i oko 200 različitih nekatalitičkih proteinskih faktora.
Molekul proteina je velika grupa aminokiselina, koja sadrži u prosjeku 100 do 500 aminokiselina. Program za sintezu proteina kože pohranjen je u molekulu deoksiribonukleinske kiseline (DNK). Molekul DNK je polimer čiji su monomeri nukleotidi. Redoslijed azotnih baza u molekulu DNK određuje slijed aminokiselina u proteinskom molekulu.
Molekul DNK ima četiri tipa azotnih baza: adenin (A), gvanin (G), citozin (C) i timin (T). Slijed od tri supstituenta (triplet) se koristi za formiranje kodona koji predstavlja jednu aminokiselinu.
Nukleinske kiseline – DNK i RNK – su vezivne komponente biosinteze proteina. DNK je odgovorna za pohranjivanje genetskih informacija, dok je RNA odgovorna za prijenos ovih informacija i implementaciju u proteinske molekule. Može se reći da je glavna funkcija DNK očuvanje genotipa, a RNK očuvanje virusnog genotipa.
U kulturi, ribosomalna RNK (rRNA) je važnija. rRNA može biti spiralizirana i sadržavati modificirane nukleotide (na primjer, 2-metil ribozu). rRNA postaje otprilike 80% ukupne količine RNK u stanicama. Drugi tip RNK u ćelijama predstavlja transfer RNK (tRNA), koja se, kao i druge vrste RNK, sintetizira u jezgru. Ovaj dio čini 10-15% ukupne količine RNK u stanicama. Identificirano je preko 60 različitih tRNA. Također, transport aminokiselina ovisi o nizu različitih tRNA. Postoji jedna specifična tRNA za kožne aminokiseline u ćelijama. Molekuli tRNA su identični. Njihova struktura ima 75-93 ribonukleida.
Aminokiselina se dodaje jakoj 3-OH grupi terminalnog mononukleotida tRNA, koja je prva predstavljena adenilnom kiselinom. tRNA ima još jedan važan dio - antikodon, pored kojeg se kompleks aminokiselina i tRNA koristi za određivanje sekvence tri nukleotida u šablonskoj RNK (kodon). Antikodon i kodon su komplementarno povezani sa vodenim spojevima.
Budući da u ćeliji nosimo informacije o eksploziji, DNK, koja je sadržana u jezgru, a sinteza proteina se dešava u citoplazmi, tada postoji pjevački posrednik koji tu informaciju prenosi u citoplazmu ćelije. Informacijski posrednik bila je glasnička RNK (mRNA). Prije mRNA nalazi se 2% ćelijskog sadržaja RNK ćelija. Pronađeni molekuli mRNA (uključuju do 5 hiljada nukleotida). mRNA također sadrži nekoliko tipova azotnih baza. Tri od njih (A, G, C) su isti kao u DNK, a četvrti je uracil.
Informacije kodirane u mRNA neophodne su za sintezu proteinske molekule koja se proizvodi na ribosomima. Sinteza mRNA u ćelijskom jezgru je vrlo brza, što zahtijeva aktivnu biosintezu proteinskih molekula. mRNA se stvara na jednom od lanaca DNK jezgra. U ovom slučaju, dvolančana struktura DNK se odmotava i DNK sinteza RNA polimeraze slijedi princip komplementarnosti:

Šema za sintezu mRNA
Princip komplementarnosti znači da je adenin na spirali DNK predstavljen uracil mRNA, timin adeninom, a gvanin citozinom. Takođe, mRNA čita informacije iz DNK.
Faza DNK - RNK, dakle, znači sintezu molekula mRNA, u kojoj je nukleotidna sekvenca komplementarna sekvenci DNK (gena). Ovaj proces se zove transkripcija. Zatim mRNA stiže do ribozoma, spajajući se sa njegovim podjedinicama. Jedna molekula mRNA istovremeno je fiksirana na više ribozoma, takozvanih polizoma. Prisustvo polisoma promoviše efikasnost i fluidnost proizvodnje mRNA.
Sinteza polipeptidnog lanceta se izvodi na mRNA šablonu. Proces prijenosa sa mRNA na protein naziva se translacija. Faza “RNA -> protein” je proces sinteze proteina, koji je usmjeren na mRNA. Dakle, prijenos informacija uvijek ide direktno preko DNK - RNK - proteina.
Proces prevođenja uključuje sljedeće faze:
Ovaj enzim učestvuje u fiksaciji prethodno aktivirane aminokiseline na pozicijama 2 ili 3 riboze preostalog tRNA nukleotida:
U ovom kompleksu, aminokiselina se transportuje do ribozoma i dolazi do sinteze proteinskog molekula. Aminoacil-tRNA sintetaza je specifična i može prepoznati i aminokiselinu i tRNA. U klinitu postoji čak 20 različitih sintetaza, u zavisnosti od broja a-aminokiselina.
2. tRNA, povezana esterskom vezom sa aminokiselinom, stiže do ribosoma i stupa u interakciju s mRNA prema tipu komplementarnosti između specifičnog tripleta mRNA nukleotida, nazvanog kodon, i njegovih komplementarnih specifičnih triplet nukleotida (antikodon) tr. . Dakle, kodon mRNA ukazuje na specifičnu fiksaciju jedne aminokiseline u peptidnom lankusu iza odgovarajućeg antikodona tRNA. Ribosom razgrađuje molekule mRNA, uzastopno čitajući sve kodone, čime se uspostavlja redoslijed distribucije svih aminokiselina koje se isporučuju na mjesto sinteze.
Sinteza proteinske molekule teče direktno od jake amino grupe do jake karboksilne grupe aminokiseline. Odredite da je aminokiselina bakalara u sintezi polipeptidne lance metionin, za koji je kodon nukleotidna sekvenca AUG mRNA.
Pokretanje sinteze polipeptida počinje fiksacijom dva tRNA antikodona iza kodona mRNA. Proces jasno uključuje energiju GTP-a, kao i učešće niza proteinskih faktora u pokretanju i peptidil transferaze.
Zahvaljujući ovom enzimu, tečnost kovalentne veze dostiže 1200 aminokiselina/komponenti/ribozoma.

Šema iniciranja sinteze polipeptida
3. Nakon stvaranja "nevezanog" dipeptida, tRNA onesposobljava ribozom i može isporučiti nove molekule aminokiselina, a mRNA se ubacuje u ribozom (polisom) za tri nukleotida. Kao rezultat kretanja (translokacije), slobodni kodon zauzima poziciju za prepoznavanje ciljne tRNA molekule. Takođe, u fazi elongacije dolazi do sukcesivnog dodavanja jedne aminokiseline polipeptidnoj lanceti u podvrsti u red kodona molekula mRNA.
Polipeptidni lancet, koji je kombinovan sa jednim tRNA molekulom, fiksiran je za veliku podjedinicu ribozoma. Dodavanje dodatne aminokiseline kože polipeptidnoj koplji je zbog interakcije amino grupe aminokiseline koja je u kompleksu sa tRNA i karboksilnom grupom peptida.
4. Završetak ili završetak sinteze polipeptidnog molekula proizvodi terminacijske kodone “bez smisla” i faktore terminacije proteina. Postoje tri kodona (UAG, UGA, UAA) koji ne kodiraju niti vezuju nijednu aminokiselinu, budući da u ćeliji nema tRNA antikodona koji su im komplementarni. Teoretski, samo jedan "bez smisla" kodon, koji proizvodi polizom tokom prolaska 5-3 mRNA, odgovoran je za sporu sintezu proteinskog molekula.
Prisustvo terminacionog kodona u bilo kojoj sekvenci mRNA znači terminaciju sinteza proteina. Kao rezultat, polizom se raspada, nevikristan mRNA se hidrolizira polinukleotid fosforilazom, a ribosomske podjedinice se pripremaju prije sinteze novog proteinskog molekula.
mRNA može više puta učestvovati u procesu biosinteze proteina. Funkcionalnost molekula mRNA varira u različitim organizmima. Može varirati od mnogo hvilina do mnogih dibova.
5. DNK kodira originalnu strukturu proteina. Stoga su proteinski molekuli sintetizirani na ribosomima još uvijek u svom konačnom stanju. Oni predstavljaju primarne polipeptide, koji se potom prepoznaju numeričkim modifikacijama (povezivanje monomera sa oligomerima, dodavanje koenzima, hemijske reakcije) koje menjaju strukturu proteina, a samim tim i njihovu aktivnost.
Sekundarne i tercijarne strukture nisu kodirane, one su označene snagama primarne strukture, a to znači da drugi oblik proteinske molekule leži u slijedu aminokiselina i mogućnostima njihove međusobne interakcije. Strukturne modifikacije proteina koji se sintetiziraju odvijaju se čak i na ribosomima ili nakon završetka sinteze kao rezultat dodavanja različitih funkcionalne grupe.
Pogledao dijagram prijenosa na prvi pogled
Možete se mijenjati u napadima i počecima. Dakle, kod virusa koji ne uništavaju DNK, informacije su pohranjene u RNK. Kada virus prodre u ćeliju, ova informacija se prenosi u DNK stanice, a ostatak potom sintetizira mRNA na kojoj se sintetiziraju virusni proteini. Ovaj proces se naziva transkripcija prolaza, a shema prijenosa u ovom slučaju će biti:
Dok je sekvenca nukleotida DNK i, prema tome, mRNA očuvana, priroda novosintetiziranog proteina ostaje nepromijenjena.
Neophodne genetske informacije za sintezu proteina mogu se predstaviti na sličan način kao i zapis ljudskog jezika, koji se sastoji od niza slova koji formiraju riječi i govor. Genetski materijal, međutim, ima više od jedne bukvalne baze (adenin, gvanin, uracil, citozin).
Genetski kod uključuje triliteralne riječi. Ako unesete u ovaj odjeljak (43) dobit ćete 64 varijante (riječi), koje su više nego dovoljne za kodiranje 20 aminokiselina. Dakle, 64 kodona formiraju genetski kod (tabela 3).
Analiza genetskog koda pokazuje da različite aminokiseline imaju različit broj kodona. Na primjer, metionin i triptofan sadrže samo jedan kodon, dok arginin, leucin i serin sadrže šest kodona. Prisustvo više kodona za jednu aminokiselinu odražava "nevinost" koda. Pa, ta ista aminokiselina može biti kodirana iza vlastitih nukleotidnih tripleta. U isto vrijeme, kožni triplet se identificira po cijeloj aminokiselini u polipeptidnoj lanceti koja se sintetiše.
Tabela 3
Genetski kod
|
nukleotida |
Drugi nukleotid |
nukleotida |
|||
Genetski kod je univerzalan i isti kod vrsta različitih nivoa razvoja (ljudi, životinje, biljke, mikroorganizmi). Univerzalnost koda pokazuje da svi živi organizmi pripadaju jednom pretku.
Druge aminokiseline (hidroksiprolin, oksilizin), na primjer, ne mijenjaju kodon i odobravaju se korištenjem dodatnih hemijske reakcije već nakon sinteze polipeptidnog lanceta. Ovaj proces se naziva posttranslacijska modifikacija i vrlo je važan za pravilno funkcioniranje proteina kože.
Neglutinozni kodoni (UAA, UAG, UGA) ne kodiraju aminokiseline, već zapravo signaliziraju završetak sinteze proteinske molekule.
Dakle, mRNA je direktni nosilac genetske informacije od jezgra do ribozoma u citoplazmi. Jedan ribosom zauzima otprilike 80 nukleotida po mRNA mjestu i sposoban je katalizirati približno 100 peptidnih veza po liniji (Severin E. S. et al., 2011).
Sintetizirani proteinski molekuli mogu biti podvrgnuti strukturnim modifikacijama u isto vrijeme kao i ribosomi ili nakon završetka sinteze kao rezultat dodavanja različitih funkcionalnih grupa. Citoplazma mRNA ima relativno kratak period. Velika količina mRNA se sintetiše i pohranjuje u neaktivnom obliku, spremna za brzu sintezu proteina. Velik dio informacija u mRNA povezan je s linearnom sekvencom nukleotida, a integritet ove sekvence je izuzetno važan. Otpad ili promjena u redoslijedu nukleotida mogu promijeniti sintezu proteina. Do danas je u ćelijama ustanovljen niz niskih inhibitora replikacije DNK (antibiotici, hemijski agensi, antivirusni lekovi). Promjena u sekvenci purinskih ili pirimidinskih baza u genu nazvana je mutacija.
Zamjena samo jednog nukleotida u kodonu (mutacija) rezultira promjenom kodiranja jedne aminokiseline u drugu. Na primjer, mutacija je povezana sa zamjenom glutaminske kiseline valinom u molekuli hemoglobina, što dovodi do sinteze hemoglobina, što uzrokuje srpastu anemiju. Danas je poznato preko 200 mutacija polipeptidnog jezgra molekula ljudskog hemoglobina. Često su mutageni spojevi (nitrosupstituti, na primjer) koji mijenjaju strukturu azotnih baza, što dovodi do promjene u prirodi komplementarnosti baza. Ultraljubičasto zračenje uzrokuje kondenzaciju viška timina sa stvaranjem timinskih dimera. Na sreću, zbog nesretnog priliva ultraljubičastog zračenja, stvorenja su bila zaštićena loptom od atmosfere ozona.
Postoje mnogi antibiotici koji se koriste u veterinarskoj praksi za inhibiciju sinteze bakterijskih proteina (linkomicin, eritromicin, hloramfenikol) čak iu fazi translacije. Kada se to dogodi, mikrobno tkivo ginekologa počinje usporavati svoj razvoj. Antibiotici kao što su tetraciklini ne utiču na sintezu ribosoma u ćelijama živih bića. Penicilini nisu direktni inhibitori sinteze proteina; njihovo djelovanje na bakterije povezano je s blokiranjem sinteze heksapeptida stanične stijenke. Treba napomenuti da se sinteza proteina odvija ne samo na ribosomima, već i na mitohondrijima. Mitohondrije obezbeđuju dodatni i nezavisni mehanizam za sintezu proteina za sopstvene potrebe, iako se svi mitohondrijski proteini ne sintetiziraju u ovim organelama. Mitohondrijska RNK postaje manje od 3% sadržaja RNK ćelije. Ribosomi mitohondrija su manje veličine i manje citoplazmatski. UGA kodon, kao terminator za sintezu proteina u citoplazmi, sintetizira se u mitohondrijima pored UGG kodona za kodiranje aminokiseline.
Proteini sintetizirani na ribosomima ne ostavljaju potpuno potpuno stanje. Oni predstavljaju primarne polipeptide, koji se zatim prepoznaju numeričkim modifikacijama (povezivanje monomera sa modifikovanim oligomerima, dodavanje koenzima, hemijske reakcije) koje modifikuju strukturu proteina i, iz iste, jogo aktivnosti.
Slika 9 iz prezentacije “Biosinteza proteina” prije nastave biologije na temu “Biosinteza proteina”
Dimenzije: 960 x 720 piksela, format: jpg. Da biste bezbedno preuzeli sliku za lekciju biologije, kliknite desnim tasterom miša na sliku i kliknite na „Sačuvaj slike kao...“. Da biste prikazali slike u lekciji, takođe možete besplatno preuzeti prezentaciju “Protein Biosynthesis.pptx” sa svim slikama u zip arhivi. Veličina arhive je 1719 KB.
Uključite svoju prezentaciju"Funkcije proteina" - Ovako se primaju signali iz vanjskog medija i informacije se prenose u ćeliju. Kada se 1 g proteina razgradi do krajnjih proizvoda, vidljivo je 17,6 kJ. Šta je renaturacija? Tip torbice: 9. Katalitički. Proces ažuriranja strukture proteina nakon denaturacije naziva se renaturacija. Pimenov A.V. Proteini su jedan od izvora energije u tijelu.
“Proteini rechovina” - Na primjer: kolagen. Autor biologije: Boldireva L. A. Vidimo 20 AK, koji će sadržavati proteine. . zadnjica: kuhano jaje. Amino kiseline - organski govor, Neorganski proteini su fibrilarni. Kharchovy proteini. . Suvi belci. Struktura proteina. Vikoristovuyutsya organizam za ruhu. Energetski proteini.
“Proteini i njihove funkcije” - Katalitička uloga. Funkcija motora. Koncept vjeverice. Hidroliza proteina se nastavlja sve dok se polipeptidne veze ne razbiju: Sažetak: Proteini proizvode krvne sudove, tetive i kosu. Koje su funkcije proteina? Hemijska snaga proteina Proteini učestvuju u formiranju ćelijskih membrana, organoida i ćelijskih membrana.
"Biosinteza proteina" - Spisak referenci. Entry 4. Zmíst. Biosinteza proteina u živim ćelijama. 7. 10. 9. Šema ruže i kuvanog klitina. 5. 6. 1. 8. 2. 3.
"Biosinteza proteina" - prevod (latinski: transfer, prevod). Transkripcija (lat. prepisivanje). Provjerite sami. Važnost proteina. Zmíst. Energija biosinteze. Uloga enzima. Sinteza polipeptidnog lanceta na ribosomima. 5. Koja je sekvenca i-RNA nukleotida zabilježena na fragmentu DNK: T-A-C-G-G-A-T-C-A-C-G-A -G-T-G-C-T A-U -G-C-G-U-A-G-U-G-C-U A-U-G-C-C-U-A-G-U.
"Biosinteza biologije proteina" - Mikola Kostjantinovič Kolcov (1872-1940). A.G. Glavna funkcija ribozoma je sinteza proteina. Centralna dogma (osnovni postulat) molekularne biologije je matrična sinteza. C. Antikodon - triplet nukleotida na vrhu tRNA. Biosinteza proteina. Nakon završetka sinteze, iRNA se razlaže na nukleotide.
Tema ima ukupno 8 prezentacija
© A.S. Spirin
BIOSINTEZA PROTEINA, SVIT RNKA.S. Spirin
Spirin Oleksandr Sergejovič– akademik, direktor Instituta za proteine Ruske akademije nauka, član Prezidijuma Ruske akademije nauka.
Otprilike u isto vrijeme, 1953. godine, D. Watson i F. Crick su otkrili princip strukturne (molekularne) organizacije molekula gena - deoksiribonukleinske kiseline (DNK). Struktura DNK dala je ključ za mehanizam preciznog stvaranja – reduplikacije – ekspresije gena. Tako je postala poznata nova nauka molekularne biologije. Formulisana je takozvana centralna dogma molekularne biologije: DNK i RNK i proteini. Smisao je u tome da se genetske informacije, zapisane u DNK, realizuju u obliku proteina, ali ne direktno, već preko dodatnog polimera - ribonukleinske kiseline (RNA), a od nje postoje nukleinske kiseline do proteina o kojima se ne može pregovarati. . Na ovaj način, DNK se sintetiše na DNK, obezbeđujući moć reduplikacije, tako da se izlazni genetski materijal stvara u generacijama; RNK se sintetiše na DNK, što rezultira ponovnim pisanjem i transkripcijom genetskih informacija u obliku numeričkih kopija RNK; Molekuli RNK su šabloni za sintezu proteina - genetske informacije se prevode u obliku polipeptidnih koplja. U posebnim varijantama, RNK se može kopirati u oblik DNK („povratna transkripcija“), a takođe i kopirati u oblik RNK (replikacija), ali protein ne može poslužiti kao šablon za nukleinske kiseline (izvještaj).
Takođe, sama DNK označava fluidnost organizama, skup proteina koji se stvara generacijama i znakove koji su s njima povezani. Biosinteza proteina je centralni proces žive materije, a nukleinske kiseline joj obezbeđuju, s jedne strane, program koji podrazumeva čitav skup i specifičnost proteina koji se sintetišu, as druge strane mehanizam za preciznu proizvodnju stvaranja ovaj program u generacijama. Stoga se život u svom trenutnom ćelijskom obliku svodi na mehanizam smanjene biosinteze proteina.
BIOSINTEZA PROTEINA
Centralna dogma molekularne biologije postulira prijenos genetskih informacija s nukleinskih kiselina na proteine i, prema tome, na komponente živog organizma. Razvoj mehanizama implementacije ovog procesa u periodu od deset godina, koji je pratio formulaciju centralne dogme, otkrio je širok spektar funkcija RNK, ne samo kao nosioca informacija iz gena. ív (DNK) na proteine i služe kao matrica za sintezu proteina.
Na sl. Slika 1 prikazuje osnovni dijagram biosinteze proteina u ćelijama. glasničku RNA(messenger RNA, messenger RNA, mRNA), koja kodira proteine, kao što je gore navedeno, samo je jedna od tri glavne klase ćelijskih RNK. Glavna masa (oko 80%) postaje druga klasa RNK - ribosomska RNK, koji stvaraju strukturni okvir i funkcionalne centre univerzalnih čestica koje sintetišu proteine – ribozoma. Sama ribosomska RNK je jedinstvena - i strukturno i funkcionalno - u formiranju ultramikroskopskih molekularnih mašina zvanih ribosomi. Ribosomi primaju genetske informacije kao mRNA molekule i, programirani od strane ostalih, proizvode proteine koji su potpuno slični ovom programu.
Proteini, da bi se sintetizirali proteini, bez informacija ili programa nisu dovoljni - potreban materijal od kojeg se mogu koristiti. Protok materijala za sintezu proteina odvija se u ribosomima uz pomoć treće klase ćelijskih RNK. RNA nosioci(Transferna RNK, transportna RNK, tRNA). Oni kovalentno vezuju - prihvataju - aminokiseline, koje su građevinski materijal za proteine, a u obliku aminoacil-tRNA nalaze se u ribosomu. U ribosomima, aminoacil-tRNA stupa u interakciju sa kodonima - kombinacijama trinukleotida - mRNA, što rezultira dekodiranjem kodona tokom procesa translacije.
Ribonukleinske kiseline
Stoga, možemo regrutirati osnovne ćelijske RNK, početni glavni proces svakodnevne žive materije je biosinteza proteina. Ce mRNA, ribosomalna RNA i tRNA. RNK se sintetizira na DNK uz pomoć enzima - RNA polimeraza, koje vrše transkripciju - prepisivanje pojedinačnih dijelova (linearnih jedinica) dvolančane DNK u oblik jednolančane RNK. Dijelovi DNK koji kodiraju ćelijske proteine se prepisuju u obliku mRNA, tako da za sintezu brojnih kopija ribosomske RNK i tRNA postoje posebni dijelovi ćelijskog genoma koji se intenzivno prepisuju bez daljeg prevođenja. í̈ proteini.
Hemijska struktura RNK.
Hemijski je RNK vrlo slična DNK. Uvredljivi govor je cijeli linearni polimer nukleotida. Monomer kože - nukleotid - je fosforilirani N-glikozid, uzrokovan viškom petougljičnog atoma - pentozila, koji nosi fosfatnu grupu na hidroksilnoj grupi petog atoma ugljika (sklopiva ugljikova veza) i dušikovu bazu na prvom atomu ugljika i (N-glikozin) Glavna hemijska razlika između DNK i RNK je u tome što RNA monomer ima ceribozu, a DNK monomer ima deoksiribozu, koja je slična ribozi, koja ima jednu hidroksilnu grupu na drugom atomu ugljika (Sl. 2).
 Mala 2. Hemijske formule viška jedan od ribonukleotida - uridila kiselina (U) i homologna yomu deoksiribonukleotid - timidilna kiselina (dT) |
Postoje dvije vrste azotnih baza u DNK i RNK: dva purina - adenin (A) i gvanin (G) - i dva pirimidina - citozin (C) i uracil (U) ili metilirani timin (T). Uracil je karakterističan za RNK monomere, a timin je karakterističan za DNK monomere, a ovo je druga vrsta RNK i DNK. Monomeri - ribonukleotidi RNK ili deoksiribonukleotidi DNK - formiraju polimerno lance kroz dodatno formiranje fosfodiestarskih mjesta između krvavih viška (između petog i trećeg atoma pentokarbonskog ugljika). Dakle, polimerni lancet nukleinske kiseline - DNK ili RNA - može se predstaviti kao linearna šećerno-fosfatna kičma sa azotnim bazama kao biološkim grupama. Makromolekularna struktura RNK. Osnovna makrostrukturna sličnost dvije vrste nukleinskih kiselina leži u činjenici da je DNK jedna subheliksa, makromolekula dva komplementarna pletena polimerna lanca, spiralno uvijena oko bočne ose (div. [, ]), a RNK je jednolančani polimer. Istovremeno, interakcija bioloških grupa - azotnih baza - jedne s jednom, kao i sa fosfatima i hidroksilima šećerno-fosfatne kičme, dovodi do toga da jednolančani RNA polimer sagorijeva na sebe i uvija se u kompaktna struktura, slična spaljivanju polipeptidnog koplja hektara proteina u kompaktnoj globuli Dakle, jedinstvene nukleotidne sekvence RNK mogu formirati jedinstvene prostorne strukture. |
Po prvi put, specifična prostorna struktura RNK je demonstrirana kada je dešifrovana atomska struktura jedne od tRNA 1974. [ , ] (slika 3). Larinks polimerne lance tRNA, koji se sastoji od 76 nukleotidnih monomera, formiran je tako da formira kompaktno globularno jezgro, iz kojeg se dvije izbočine direktno trljaju jedna uz drugu. To su kratke podlančane spirale na jezgru DNK, ali su organizirane kroz interakciju dijelova istog lanca RNK. Jedan od njih djeluje kao akceptor aminokiselina i učestvuje u sintezi polipeptidnog proteina na ribosomu, a drugi se koristi za komplementarnu interakciju sa tripletom (kodonom) mRNA koji kodira u istom ribosomu. Samo je ova struktura dizajnirana za specifičnu interakciju s proteinom-enzimom koji vezuje aminokiselinu za tRNA, i ribozomom tokom translacije, tako da je oni specifično "prepoznaju".

Mala 3. Atomski (ljevoruki) i skeletni (desnoruki) modeli tRNK fenilalanina kvasca
Infuzija izolovanih ribosomskih RNK je pružila napredno rešenje za formiranje kompaktnih, specifičnih struktura još dužih linearnih polimera ovog tipa. Ribosom se sastoji od dva nejednaka dijela - velike i male ribosomske podjedinice (podjedinice). Podčestica kože sastoji se od jedne visokopolimerne RNK i niza različitih ribosomskih proteina. Broj ribosomskih RNK je čak značajan: na primjer, RNK male podjedinice bakterijskog ribosoma sadrži preko 1500 nukleotida, a RNK velike podjedinice sadrži približno 3000 nukleotida. Kod ljudi, uključujući ljude, ciRNA je još veća - oko 1900 nukleotida i preko 5000 nukleotida u malim i velikim podjedinicama vrste.
Pokazalo se da su izolirane ribosomske RNK poboljšane od svojih proteinskih partnera i izolirane od čist izgled, same strukture spontano kolabiraju u kompaktnu strukturu, sličnu po veličini i obliku kao ribosomske podjedinice]. Oblik velikih i malih subčestica varira, a oblik velike i male ribosomske RNK varira u skladu s tim (slika 4). Tako se linearni lanci ribosomske RNK samoorganiziraju u specifične prostorne strukture, koje određuju veličinu, oblik i, možda, unutarnju strukturu ribosomskih podjedinica, a time i cijelog ribosoma.
Minor RNA. U svijetu je inokulacija komponenti živih stanica i drugih frakcija ukupne ćelijske RNK otkrila da se tri glavne vrste RNK na desnoj strani ne mogu razdvojiti. Pokazalo se da u prirodi ne postoje druge vrste RNK. To je ono što nazivamo “mala RNK”, koja sadrži do 300 nukleotida, često s nepoznatim funkcijama. U pravilu su povezani s jednim ili više proteina i predstavljeni su u stanicama kao ribonukleoproteini - "mali RNP".
Male RNA prisutne su u svim dijelovima ćelije, uključujući citoplazmu, jezgro, jezgro i mitohondrije. Većina ovih malih RNP-ova, čije funkcije učestvuju u mehanizmima post-transkripcione obrade glavnih tipova RNK (obrada RNK) - konverzija prekursora mRNA u zrelu mRNA (splicing), uređivanje mRNA, biogeneza tRNA, doziranje ribosomska RNK. Jedan od najzastupljenijih tipova malih RNP-ova (SRP) u ćelijama igra ključnu ulogu u transportu proteina koji se sintetiziraju kroz ćelijsku membranu. Postoje vrste malih RNK koje obavljaju regulatorne funkcije u translaciji. Posebna mala RNK ulazi u skladište najvažnijeg enzima odgovornog za podršku reduplikaciji DNK u generacijama ćelija - telomeraze. Treba reći da njihove molekularne dimenzije mogu biti jednake dimenzijama ćelijskih globularnih proteina. Dakle, postaje sve jasnije da funkcionisanje žive ćelije nije određeno samo raznovrsnošću proteina koji se u njoj sintetiziraju, već i prisustvom bogatog skupa raznovrsnih RNK, od kojih su male RNK značajne. kompaktnost i veličina proteina.
Ribozyme. Sav aktivni život vođen je razmjenom supstanci – metabolizmom, a sve biohemijske reakcije metabolizma se odvijaju sa potrebnim tečnostima za obezbeđivanje života, samo uz visokoefikasne specifične katalizatore, uz koje stvaramo evoluciju. Već dugi niz decenija, biohemija je bila uvjerena da se biološku katalizu uvijek izvode proteini tzv. enzimi, ili enzimi. I os y 1982-1983 pp. Pokazalo se da u prirodi postoje vrste RNK koje, poput proteina, imaju visoko specifičnu katalitičku aktivnost [,]. Ovi RNA katalizatori su nazvani ribozimi. Znanju o krivici proteina u katalizi biohemijskih reakcija došao je kraj.
Danas se ribozom takođe smatra ribozimom. Istina, svi očigledni eksperimentalni podaci ukazuju na to da sintezu polipeptidnog proteina u ribosomu katalizira ribosomalna RNK, a ne ribosomalni proteini. Identifikovana je katalitička komponenta velike ribosomalne RNK, koja je odgovorna za katalizu transpeptidacione reakcije, što takođe doprinosi rastu proteinskog polipeptida tokom translacije.
Što se tiče replikacije virusne DNK, njen mehanizam se malo razlikuje od replikacije genetskog materijala - DNK - same ćelije. U nekim virusnim RNK ostvaruju se procesi koji su potisnuti ili cijeli dan u normalnim stanicama, gdje se sva RNK sintetizira samo na DNK kao na matriksu. Kada se zarazi RNA virusima, situacija može biti dvostruka. U nekim slučajevima, DNK se sintetiše na virusnoj RNK, kao na matriksu („povratna transkripcija“), a na ovu DNK se transkribiraju brojne kopije virusne RNK. U drugim slučajevima, koji su za nas relevantni, komplementarni lanac RNK se sintetiše na virusnoj RNK, koja služi kao šablon za sintezu - replikaciju - novih kopija virusne RNK. Na taj način, prilikom infekcije RNA virusima, ostvaruje se princip da RNK određuje stvaranje njene strukture, kao i DNK.
Multifunkcionalnost RNK. Pretpostavka i opće znanje o funkcijama RNK omogućavaju nam da govorimo o jedinstvenoj funkcionalnosti ovog polimera u živoj prirodi. Moguće je uvesti trenutnu promjenu u osnovne funkcije RNK.
Genetska replikativna funkcija: strukturna sposobnost kopiranja (replikacije) linearnih nukleotidnih sekvenci kroz komplementarne sekvence. Funkcija se realizuje tokom virusnih infekcija i slična je glavnoj funkciji DNK u živim ćelijskim organizmima - reduplikaciji genetskog materijala.Uopšteno govoreći, RNK stoji pred nama kao čudesan polimer, za koji se čini da vreme za evoluciju Univerzuma, niti intelekt Stvoritelja, ne bi učinili svoje. Kako se ispostavilo, RNK je dizajnirana da kombinuje funkcije oba fundamentalno važna polimera - DNK i proteina. Nije iznenađujuće što je prehrana došla prije nauke: zašto osjećaj krivice i samodovoljno postojanje RNK svjetlosti ne bi mogao prenijeti izgled života u njegovom trenutnom obliku DNK-proteina?Funkcija kodiranja: programiranje sinteze proteina linearnim sekvencama nukleotida. To je upravo funkcija koju ima DNK. I u DNK i u RNK isti triplet nukleotida kodira 20 aminokiselina proteina, a sekvenca tripleta u sekvenci nukleinske kiseline sadrži program za sekvencijalni raspored 20 vrsta aminokiselina u polipeptidnoj sekvenci južne vjeverice.
Funkcija stvaranja strukture: formiranje jedinstvenih trivijalnih struktura. Kompaktno presavijene male RNK molekule su u osnovi slične trivijalnim strukturama globularnih proteina, a veće RNK molekule mogu stvoriti veće biološke dijelove ili njihova jezgra.
Funkcija prepoznavanja: visoko specifične interakcije sa drugim makromolekulama (uključujući proteine i druge RNK) i sa malim ligandima. Ova funkcija je vjerovatno glavna u proteinima. Zasniva se na sposobnosti polimera da formira jedinstvenu strukturu i formira specifične trivijalne strukture. Funkcija prepoznavanja je osnova specifične katalize.
Katalitička funkcija: specifična kataliza kemijskih reakcija ribozimima. Ova funkcija je slična enzimskoj funkciji enzimskih proteina.
LIFE HAVEN
Oparinova teorija protein-koacervata. Možda je, prema nauci, dobro osmišljenu teoriju življenja duž abiogenog puta predložio biohemičar A.I. Oparin je rođen 20-ih godina prošlog vijeka [,]. Teorija se zasnivala na otkriću da je sve počelo sa proteinima i na mogućnosti u ranim umovima spontane hemijske sinteze proteinskih monomera - aminokiselina - i polimera sličnih proteinima (polipeptida) pomoću abiogenih supstanci. Lyakh. Objavljivanje teorije podstaklo je numeričke eksperimente u brojnim laboratorijama širom svijeta, koji su pokazali realnost takve sinteze u individualnim umovima. Švidkova teorija postala je široko prihvaćena i izuzetno popularna.
Njegov glavni postulat bio je da su se čestice slične proteinima spontano pojavile u originalnom „bujonu“ i da su kombinovane „u koacervatne kapi - koloidni sistemi (soli) ojačani vodom koji plutaju u razrijeđenijem vodenom rastvoru. promijeniti um krivice organizama - jačanje dovkilla, i kompartmentalizacija S obzirom da je nekoliko čestica koacervata nalik na proteine moglo imati katalitičku aktivnost, bilo je moguće da se u sintezi pjega dogode biokemijske reakcije - slični proizvodi. Postoji asimilacija, što znači rast koacervata uz daljnju dezintegraciju na dijelove - reprodukcija. Koacervat, koji asimilira, raste i razmnožava se deljenjem, smatra se prototipom živih ćelija (slika 5).
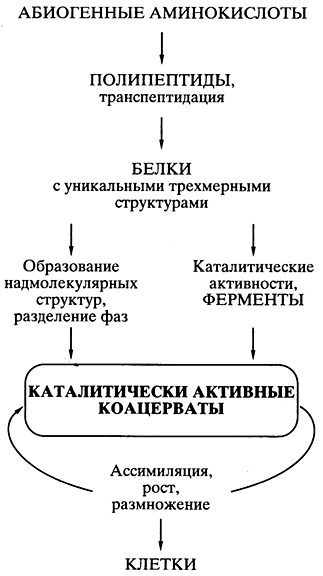
Mala 5.Šematski iskazi o putevima života
na osnovu proteinsko-koacervatne teorije A.I. Oparina
Sve je bilo dobro osmišljeno i naučno utemeljeno u teoriji, osim jednog problema, koji je odavno spljošten očima svih lažnjaka u Galušinom životu. Pošto su spontano, u nizu nasumičnih sinteza bez šablona u koacervatu, jedna po jedna nastala strukture proteinskih molekula (na primjer, efikasni katalizatori koji bi osigurali superiornost ovog koacervata u njegovom rastu i razmnožavanju), kako bi onda copy yuvatsya za proširenje sredine koacervata, a još više za prijenos na koacervate - Teorija Pokazalo se neosnovanim isticati problem preciznog stvaranja - u sredini koacervata i u generacijama - pojedinačnih učinkovitih proteinskih struktura koje iznenada pojavio.
Lagana RNK kao nasljednik svakodnevni život. Akumulirano znanje o genetski kod, nukleinske kiseline i biosinteza proteina doveli su do konsolidacije fundamentalno nove ideje o TOM-u, da sve nije počelo od proteina, već od RNK [-]. Nukleinske kiseline su jedinstvena vrsta bioloških polimera, čija makromolekularna struktura, zbog principa komplementarnosti u sintezi novih Lancsugova (izvještaji), osigurava mogućnost kopiranja monomernih traka vlažnog linearnog niza, drugim riječima, mogućnost stvaranja (replikacije) polimera, njegove mikrostrukture. Stoga, samo nukleinske kiseline, a ne proteini, mogu biti genetski materijal, stvarajući molekule koji ponavljaju svoju specifičnu mikrostrukturu u generacijama.
Na niskim nivoima, sama RNK, a ne DNK, može biti primarni genetski materijal.
Prema prvom a u hemijskoj sintezi i biohemijskim reakcijama ribonukleotidi se prenose u deoksiribonukleotide; Deoksiribonukleotidi su produkti modifikacije ribonukleotida (div. sl. 2).
na drugačiji način, U modernim, univerzalnim procesima živog metabolizma, široko su zastupljeni sami ribonukleotidi, a ne dezoksiribonukleotidi, uključujući i glavne nosioce energije tipa ribonukleozid polifosfata (ATP).
na trećem, RNA Replykatsya je moguća bez ikakve DNK, a mehanizam je reduciran Vijećem Vijeća Žive Vimage, RNA-Strauvka u INITSIATSIC SNITY LANTSYA DNK.
Na četvrtom Uz sve iste matrične i genetske funkcije koje su stvorene DNK i RNK, one također imaju niske funkcije koje kontroliraju proteine, uključujući katalizu kemijskih reakcija. Dakle, postoje svi razlozi da se DNK posmatra kao kasniji evolucijski dodatak – kao modifikacija RNK, specijalizovana za specifičnu funkciju stvaranja i očuvanja jedinstvenih kopija gena u genomu skladišta ćelija bez ikakvog posredničkog učešća u biosintezi proteina.
Nakon što su otkrivene katalitički aktivne RNK, na razvoj ideje o primatu RNK u svakodnevnom životu snažno je utjecalo i koncept je formuliran. samodovoljan svijet RNK, otpuhavši svakodnevni život [,]. Mozhlivov dijagram vinifikacije RNK predstavljen je na Sl. 6.
Abiogena sinteza ribonukleotida i njihovo kovalentno formiranje u oligomere i polimere tipa RNK mogla se desiti u približno istim umovima i u istim hemijskim uslovima koji su pretpostavljeni za stvaranje aminokiselina i polipeptida. Nedavno O.B. Chetverin i spivrobytniki (Institut za proteine RAS) eksperimentalno su pokazali da se primarne komponente poliribonukleotida (RNA) u osnovnoj vodenoj sredini stvaraju prije spontane rekombinacije, tako da dolazi do zamjene fragmenata.To je način transesterifikacije. Razmjena kratkih fragmenata lanceta za duge dovodi do redukcije poliribonukleotida (RNA), a sama takva rekombinacija je u skladu sa strukturnom raznolikošću ovih molekula. Među njima bi mogle biti katalitički aktivne RNK molekule.
Rijetka pojava pojedinačnih RNA molekula, koji katalizuju polimerizaciju ribonukleotida ili spajanje oligonukleotida na komplementarnoj RNA matrici [ ], značilo je uspostavljanje mehanizma replikacije ikacione RNA. Replikacija RNA katalizatora (ribozima) sama po sebi uzrokuje malo štete samoreplicirajućim RNA populacijama. Proizvodnja vlastitih kopija RNK se umnožava. Neizbježne promjene u kopiranju (mutaciji) i rekombinaciji u RNK populacijama, koje se samorepliciraju, stvarale su sve veću raznolikost u ovom svijetu. Dakle, prijenos drevnog svijeta RNK - tse "samodovoljan biološki svijet u kojem su molekule RNK funkcionirale kao genetski materijal i kao katalizatori slični enzimima" .
Krivica za biosintezu proteina. Dalje, na osnovu lake RNK, postoji mali napredak u formiranju mehanizama biosinteze proteina, pojavljivanju različitih proteina sa urušenom strukturom i snagom, razdvajanju sistema biosinteze proteina i regrutaciji proteina i, možda, u obliku koacervati postoji evolucija preostale kliniformne strukture - živi klini (div. sl. 6) . ).
Problem prelaska sa drevne RNK svjetlosti na sadašnju svjetlost koja sintetiše proteine je najvažnija tačka za vrlo teorijsko rješenje. Obilje sinteze polu-peptida, ne pre-magino, virishenni problemi, okolka ne gleda na isti specifični šešir, jak sinteza mig Boti, il od RNA PID Genity Control. Genetička kontrola sinteze polipeptida i proteina razvija se nezavisno od primarne abiogene sinteze, na svoj način, uz regulaciju već postojeće lake RNK. Literatura je sugerirala brojne hipoteze o sličnosti svakodnevnog mehanizma biosinteze proteina u laganoj RNK, ali se možda one mogu posmatrati kao promišljene do detalja i nepredviđene sa stanovišta fizičko-hemijskih mogućnosti. Predstavljam svoju verziju procesa evolucije i specijalizacije RNK, koji dovodi do uništenja aparata za biosintezu proteina (malih 7), ali ne pretenduje da je završen.
Predlaže se hipotetička shema koja kombinuje dva bitna momenta koja se čine važnima.
Prema prvom Pretpostavlja se da se abiogeno sintetisani oligoribonukleotidi aktivno rekombinuju kroz dodatni mehanizam spontane neenzimske transesterifikacije, što dovodi do stvaranja aktivnih RNA lanaca i dovodi do njihove raznolikosti. Upravo ovaj put u populaciji oligonukleotida i polinukleotida mogao bi uključivati i katalitički aktivne tipove RNK (ribozime) i druge tipove RNK sa specijalizovanim funkcijama (slika 7). Štaviše, neenzimska rekombinacija oligonukleotida koji se komplementarno vezuju za polinukleotidni matriks mogla bi osigurati umrežavanje (spajanje) fragmenata komplementarnih matriksu u jednu lancetu. Na ovaj način, umjesto da se katalizira polimerizacijom mononukleotida, moglo bi doći do primarnog kopiranja (replikacije) RNK. Očigledno, ako je utvrđeno da ribozimi imaju nisku aktivnost polimeraze, onda je efikasnost (tačnost, fluidnost i produktivnost) kopirana na komplementarnu. Matrice se smanjuju i rastu.

Mala 7.Šema evolucije i specijalizacije molekula RNK
u procesu tranzicije iz antičkog svijeta RNK u trenutnom svetu
genetski određena biosinteza proteina
Ostalo Važna stvar u mojoj verziji je da se prvi aparat za biosintezu proteina pojavio sa rasporedom nekoliko tipova specijalizovane RNK pre pojave aparata za enzimsku (polimerazu) replikaciju genetskog materijala - RNK i DNK. Ovaj primarni aparat uključuje katalitički aktivnu proribozomalnu RNK, koja ima nisku aktivnost peptidil transferaze; skup pro-tRNA koje specifično vezuju aminokiseline ili kratke peptide; Ostala proribozomalna RNK interaguje istovremeno sa katalitičkom proribozomalnom RNK, pro-mRNA i pro-tRNA (div. Slika 7). Takav sistem bi već mogao sintetizirati polipeptidne lance putem katalizirane reakcije transpeptidacije. Među ostalim katalitički aktivnim proteinima - primarnim enzimima (enzimima) - pojavili su se proteini koji kataliziraju polimerizaciju nukleotida - replikaza, odnosno NK polimeraza.
Međutim, moguće je da hipoteza o u antički svijet RNK, kao prethodnica svakodnevnom živom svijetu, ne može dobiti dovoljno uvjeta za rješavanje glavnog problema - naučno prihvatljivog opisa mehanizma tranzicije sa RNK i replikacije u biosintezu proteina. Dodata je i detaljno osmišljena alternativna hipoteza AD. Altstein (Institut za biologiju gena Ruske akademije nauka), koji postulira da su replikacija genetskog materijala i njegova translacija - sinteza proteina - evoluirali i evoluirali istovremeno i konzumirali, počevši od međusobne interakcije abiogena ali sinteze oligonukleotida i aminoacila -nukleotidilati - miješani anhidridi. Kozak se već približava... ( "A Shahrazad je uhvaćen ujutro, i ona je dobila dozvolu za promociju".)
Književnost
1. Watson J.D., Crick F.H.C. Molekularna struktura nukleinskih kiselina // Priroda. 1953. V. 171. P. 738-740. 2. Watson J.D., Crick F.H.C. Genetske implikacije strukture nukleinske kiseline deoksiriboze // Nature 1953 V. 171. P. 964-967. 3. Spirin A.S. Dnevna biologija i biološka sigurnost // Bilten Ruske akademije nauka. 1997. br. 7. 4. Spirin A.S. O makromolekularnoj strukturi prirodnih visokopolimernih ribonukleinskih kemikalija // Journal of Molecular Biology. 1960. V. 2. P. 436-446. 5. Kirn S.H., Suddath F.L., Quigley G.J. ta in. Trodimenzionalna tercijarna struktura RNK prijenosa fenilalanina kvasca // Nauka. 1974. V. 185. P. 435-40. 6. Robertas J.D., Ladner J.E., Finch J.T. ta in. Struktura tRNA fenilalanina kvasca pri rezoluciji 3 A // Nature. 1974. V. 250. P. 546-551. 7. Vasiliev V.D., Serdyuk I.N., Gudkov A.T., SPIRin A.S. Samoorganizacija ribosomske RNK // Struktura, funkcija i genetika ribosoma / Eds. Hardesty B. i Kramer G. New York: Springer-Verlag, 1986. P. 129-142. 8. Baserga S.J., Steitz J.A. Raznolik svijet malih ribo-nukleoproteina // The RNA World / Eds. Gesteland R.F. í Atkins J.F. New York: Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1993, str. 359-381. 9. Kruger K., Grabowski PJ., Zaug AJ. ta in. Samo-spajajuća RNA: autoekscizija i autociklizacija ribosomske RNK intervenirajuće sekvence Tetrahimena// Cell. 1982. V. 31. P. 147-157. 10. Guerrier-Takada S., Gardiner K., Marsh T. et al. RNA dio ribonukleaze P je katalitička podjedinica enzima // Cell. 1983. V. 35. P. 849-857. 11. Oparin A.I.Šetnja života. M: Moskovski robotnik, 1924. 12. Oparin A.I. Krivica za život Zemlje (3 vrste). M: Ogranak Akademije nauka SRSR, 1957. 13. Woese S. Evolucija genetskog koda // The Genetic Code. New York: Harper & Row, 1967. str. 179-195. 14. Crick F.H.C. porijeklo genetskog koda // Journal of Molecular Biology. 1968. V. 38. P. 367-379. 15. Orgel L.E. Evolucija genetskog aparata // Journal of Molecular Biology. 1968. V. 38. P. 381-393. 16. Gilbert W. RNK svijet // Priroda. 1986. V 319, str. 618. 17. Joyce G.F., Orgel L.E. Izgledi za razumijevanje porijekla RNA svijeta // The RNA World / Eds. Gesteland R.F. í Atkins J.F. New York: Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1993. P 1-25. 18. Četverina H.V., Demidenko A.A., Ugarov V.I., Četverin A.B. Spontane reorganizacije u RNA sekvencama // FEBS Letters. 1999. V. 450. P. 89-94. 19. Bartel D.P., Szostak J.W. Izolacija novih ribozima iz velikog skupa nasumičnih sekvenci // Nauka. 1993. V. 261. P. 1411-1418. 20. Ekland E.H., Bartel D.P. RNA katalizirana RNA polimerizacija vikorističkih i nukleozid trifosfata // Priroda. 1996. V. 382. P. 373-376. 21. Orgel L.E. porijeklo života - Pregled činjenica i nagađanja // Trends in Biochemical Sciences. 1998. V. 23. str. 491-495. 22. Altshtein A.D. Sličnost genetskog sistema: hipoteza progena // Molekularna biologija. 1987. T. 21. str. 309-322.








